Balancing Act: Strategic Management of NADPH and ATP Cofactors in Engineered Metabolic Pathways for Biomedical Applications
This comprehensive review addresses the critical challenge of maintaining NADPH and ATP cofactor balance in engineered metabolic pathways, a fundamental requirement for efficient bioproduction in microbial cell factories and therapeutic...
Balancing Act: Strategic Management of NADPH and ATP Cofactors in Engineered Metabolic Pathways for Biomedical Applications
Abstract
This comprehensive review addresses the critical challenge of maintaining NADPH and ATP cofactor balance in engineered metabolic pathways, a fundamental requirement for efficient bioproduction in microbial cell factories and therapeutic development. We explore the foundational principles of cofactor physiology, examine cutting-edge engineering strategies including computational modeling, protein engineering, and synthetic biochemistry modules. The article provides systematic troubleshooting approaches for resolving cofactor imbalance and presents advanced validation techniques using genetically encoded biosensors and flux analysis. Designed for researchers, scientists, and drug development professionals, this resource synthesizes recent advances to guide the rational design of optimized metabolic systems for pharmaceutical production and biomedical innovation.
The Essential Roles of NADPH and ATP: Understanding Cofactor Physiology in Cellular Metabolism
Fundamental Cofactor Functions & Common Experimental Challenges
FAQ: What are the primary cellular roles of NADPH and ATP?
NADPH and ATP are essential cofactors with distinct but interconnected roles in cellular metabolism. NADPH serves primarily as a reducing agent, providing the reducing power (electrons) for anabolic biosynthesis and combating oxidative stress. Its key functions include de novo synthesis of fatty acids, cholesterol, amino acids, and nucleotides, as well as maintaining the cellular antioxidant defense system by regenerating reduced glutathione [1] [2]. In contrast, ATP functions as the universal energy currency, coupling metabolic pathways that release energy with those that require it. It provides the necessary chemical energy for biosynthesis, active transport, and cell motility by donating its high-energy phosphate group [3] [4].
FAQ: What are the most common symptoms of cofactor imbalance in an engineered pathway?
Researchers may observe several tell-tale signs when NADPH and ATP are not adequately balanced:
- Suboptimal Product Titers: The final yield of the target compound is lower than predicted by pathway flux analysis, even when precursor supply seems sufficient [3].
- Accumulation of Metabolic Intermediates: Pathway intermediates may build up due to bottlenecks at cofactor-dependent enzymatic steps.
- Reduced Cellular Growth: Cofactor imbalances can starve primary metabolism, leading to poor biomass accumulation, as seen when glucose-6-phosphate isomerase was inactivated to increase NADPH, which instead reduced cell growth [3].
- Metabolic Rearrangements: Cells may activate compensatory pathways, such as inducing a "futile cycle" of fatty acid synthesis and degradation to convert NADPH into respirable FADH2 when NADH is limited [5].
Troubleshooting Cofactor Imbalances: A Practical Guide
Troubleshooting Guide: My pathway requires significant NADPH, and its absence is a bottleneck. How can I increase NADPH availability?
Enhancing NADPH supply is a common strategy in metabolic engineering. The following table summarizes the key approaches.
Table 1: Strategies for Engineering NADPH Supply in Microbial Hosts
| Strategy | Method | Example Host | Key Enzyme(s) Targeted | Experimental Outcome |
|---|---|---|---|---|
| Enhance Oxidative PPP | Overexpress key enzymes in the oxidative branch of the pentose phosphate pathway. | Pichia pastoris | Glucose-6-phosphate dehydrogenase (ZWF1), 6-phosphogluconolactonase (SOL3) [3] | Combined overexpression increased α-farnesene production by ~12.9% [3]. |
| Introduce Heterologous Enzymes | Express a NADH kinase to convert NADH to NADPH. | Pichia pastoris | POS5 from S. cerevisiae (cPOS5) [3] | Low-intensity expression of cPOS5 aided α-farnesene production [3]. |
| Modulate Cofactor-Consuming Reactions | Downregulate or knock out non-essential NADPH-consuming reactions. | E. coli | NADPH-dependent aldehyde reductase (YahK) [4] | Repression of yahK increased 4HPAA production by 67.1% [4]. |
| Activate Alternative NADPH Sources | Leverage cytosolic or mitochondrial pathways. | Mammalian Cells / Yeast | Cytosolic/mitochondrial Isocitrate Dehydrogenase (IDH1/IDH2), Malic Enzyme (ME1/ME3) [1] [2] | Provides NADPH in compartments outside the cytosol; important for lipid synthesis and redox defense [1]. |
Troubleshooting Guide: My biosynthetic pathway is highly ATP-intensive. What are effective ways to alleviate ATP limitation?
ATP demand must be met to prevent stalling energy-intensive pathways.
- Overexpress Enzymes in ATP Regeneration Pathways: In P. pastoris, overexpressing adenine phosphoribosyltransferase (APRT), a key enzyme in the salvage pathway for AMP synthesis, helped increase ATP availability and boosted α-farnesene production [3].
- Downregulate Competitive ATP-Consuming Processes: A CRISPRi screen in E. coli identified 19 ATP-consuming enzyme genes whose repression improved product yield. For example, repressing the ATP-dependent transport gene fecE was beneficial [4]. This strategy redirects ATP from non-essential functions toward the desired pathway.
- Reduce Metabolic Shunts that Waste NADH: In P. pastoris, inactivating glycerol-3-phosphate dehydrogenase (GPD1) blocks the conversion of dihydroxyacetone phosphate to glycerol. This prevents NADH consumption in this shunt pathway, making more NADH available for oxidative phosphorylation and subsequent ATP synthesis [3].
Experimental Protocols for Diagnosing and Correcting Imbalance
Protocol 1: Rapid Assessment of Cofactor Competition Using CRISPRi Screening
This protocol is adapted from a study that developed a "Cofactor Engineering based on CRISPRi Screening (CECRiS)" strategy in E. coli [4].
- Strain Construction: Start with a base strain producing your target compound. Introduce a dCas9 protein and a library of sgRNAs targeting all known NADPH-consuming or ATP-consuming enzyme-encoding genes in the host genome.
- Cultivation and Screening: Cultivate the pooled library in a high-throughput format (e.g., 96-well deep plates) under production conditions.
- Product Analysis: Quantify the titer of your desired product in each culture.
- Hit Identification: Identify strains with improved product yields. The sgRNAs in these strains target cofactor-consuming genes whose repression liberates NADPH or ATP for your pathway.
- Validation: Construct dedicated knockout or knockdown strains for the top hit genes to confirm the phenotype.
Table 2: Key Research Reagent Solutions for Cofactor Engineering
| Reagent / Tool | Function / Application | Example Use Case |
|---|---|---|
| Genetically Encoded Biosensors (e.g., iNAP, SoNar) | Real-time, live-cell monitoring of NADPH/NADP+ or NADH/NAD+ ratios [2]. | Visualizing dynamic changes in cofactor balance in response to genetic modifications or stress. |
| CRISPRi/dCas9 System | Targeted repression of specific genes without knockout [4]. | Systematically identifying and downregulating competitive NADPH or ATP consumers, as in the CECRiS strategy. |
| Heterologous Enzymes (e.g., POS5, Transhydrogenases) | Provide alternative routes for cofactor regeneration or interconversion [3]. | Expressing NADH kinase (POS5) to convert NADH into NADPH. |
| Enzymatic Cycling Assays & LC-MS | Accurate, absolute quantification of NADPH, NADH, ATP, ADP, etc., from cell lysates [2]. | Precisely measuring intracellular cofactor pools and their ratios in different engineered strains. |
Protocol 2: Rational Engineering of NADPH and ATP Regeneration Pathways
This protocol is based on the successful multi-step engineering of Pichia pastoris for α-farnesene production [3].
- Baseline Characterization: Measure the NADPH/NADP+ and ATP/ADP ratios in your baseline production strain during the fermentation process.
- Engineer NADPH Supply:
- Modify the Pentose Phosphate Pathway (PPP): Overexpress key oxidative PPP genes like ZWF1 and SOL3. Note: Avoid complete disruption of glycolysis, as seen when PGI was inactivated, which reduced cell growth [3].
- Introduce a NADH Kinase: Integrate a gene like cPOS5 from S. cerevisiae under a tunable promoter. Fine-tune expression to avoid excessive NADH drainage.
- Engineer ATP Supply:
- Amplify the AMP Pool: Overexpress enzymes in the adenosine salvage pathway, such as APRT, to enhance the precursor pool for ATP synthesis.
- Conserve Reducing Equivalents: Inactivate shunts like the glycerol pathway by deleting GPD1 to make more NADH available for respiratory ATP generation.
- Fermentation and Validation: Ferment the final engineered strain and quantify both the target product titer and the intracellular cofactor levels to validate the improvement.
Visualizing Cofactor Balance and Engineering Strategies
The following diagram illustrates the core concepts of cofactor balancing between energy generation, redox power, and biosynthesis, and how engineering interventions can optimize this balance.
Diagram: Strategies for Balancing Cofactor Metabolism. The diagram shows how engineering interventions (red for enhancement, green for repression) can be applied to key nodes in NADPH and ATP metabolism to redirect flux toward desired biosynthetic pathways. The diagram is conceptual and does not represent a complete metabolic network.
NADPH/NADP+ as Central Redox Couples in Anabolic Processes and Cellular Defense
Troubleshooting Guide: Common NADPH Homeostasis Issues in Engineered Systems
Problem 1: Insufficient NADPH Supply Limiting Product Yields
- Symptoms: Low titers of target compounds (e.g., 5-Methyltetrahydrofolate, fatty acids, terpenoids); accumulation of oxidized glutathione (GSSG); increased cellular oxidative stress.
- Underlying Cause: The demand for NADPH in your engineered pathway exceeds the cell's native regeneration capacity. The anabolic demand for NADPH is coupled to the rate of biomass formation and the requirements of introduced pathways [6].
- Solutions:
- Amplify Native NADPH Pathways: Overexpress key enzymes from the oxidative Pentose Phosphate Pathway (PPP), such as Glucose-6-phosphate dehydrogenase (G6PD) and 6-phosphogluconate dehydrogenase (PGD) [7] [8].
- Engineer Cofactor Specificity: Switch the cofactor preference of native enzymes from NAD(H) to NADP(H). A global candidate is glyceraldehyde-3-phosphate dehydrogenase; this can be achieved by replacing conserved glutamate or aspartate with serine in the loop region [9].
- Introduce Transhydrogenases: Overexpress membrane-bound (pntAB) or soluble (sthA) transhydrogenases to convert NADH to NADPH, balancing the cofactor pool [8] [6].
- Utilize Formate Dehydrogenase: Express formate dehydrogenases (e.g., from Candida boidinii) to regenerate NAD(P)H from the oxidation of inexpensive sodium formate [8].
Problem 2: NADPH/NADP+ Imbalance Disrupting Redox State
- Symptoms: Reduced cell growth, metabolic hysteresis, failure to maintain reduced glutathione (GSH) levels, high ROS-induced cytotoxicity.
- Underlying Cause: The high NADPH/NADP+ ratio is not maintained, compromising the cell's antioxidant defense system. The NADPH system is essential for regenerating reduced glutathione and thioredoxin [7] [1].
- Solutions:
- Modulate Carbon Flux: Reroute carbon flux into the PPP. This can be done by knocking out glycolytic genes like 6-phosphofructokinase 1 (pfkA) to increase the pool of glucose-6-phosphate available for the PPP [8].
- Enhance NADPH Generation from TCA Cycle: Overexpress cytosolic or mitochondrial isoforms of NADP-dependent isocitrate dehydrogenase (IDH1/IDH2) or malic enzyme (ME1/ME3) to generate NADPH from mitochondrial TCA-cycle intermediates [7] [1].
- Overexpress NAD Kinase (NADK): Increase the total pool of NADP+ available for reduction to NADPH by amplifying the activity of NADK, which phosphorylates NAD+ to NADP+ [7].
Problem 3: Inefficient Coupling of NADPH Regeneration to Biosynthesis under Non-Growing Conditions
- Symptoms: Product synthesis halts when growth ceases (e.g., under nitrogen limitation); carbon flux is not efficiently redirected from central metabolism to production pathways.
- Underlying Cause: The metabolic network lacks a mandatory link between product formation and redox balance during stationary or production phases.
- Solutions:
- Design Coupled Pathways: Implement pathways where product formation is essential for recycling NADP+ back to NADPH. For example, in an acetol-producing E. coli strain, the NADPH-dependent acetol biosynthesis pathway becomes crucial for maintaining the NADPH/NADP+ balance during nitrogen-limited, non-growing conditions [10].
- Use Nutrient-Limited Conditions: Strategically use nutrient limitation (e.g., nitrogen) to uncouple growth from production and force flux re-routing towards the NADPH-consuming product pathway [10].
Frequently Asked Questions (FAQs)
Q1: What are the primary metabolic pathways for NADPH generation, and how do I choose which one to engineer?
The primary pathways and their key enzymes are summarized in the table below [7] [1].
Table 1: Major NADPH-Generating Pathways and Their Features
| Pathway | Key Enzyme(s) | Compartment | Notes and Engineering Considerations |
|---|---|---|---|
| Pentose Phosphate Pathway (PPP) | G6PD, PGD | Cytosol | The largest contributor to cytosolic NADPH [11]. Ideal for boosting reducing power for cytosolic anabolism. Flux is feedback-inhibited by NADPH [11]. |
| Isocitrate Dehydrogenase | IDH1 (cytosol), IDH2 (mitochondria) | Both | Links TCA cycle to NADPH production. Requires citrate export from mitochondria for cytosolic IDH1 [1]. |
| Malic Enzyme | ME1 (cytosol), ME3 (mitochondria) | Both | Converts malate to pyruvate, generating NADPH. Part of a cycle that also produces cytosolic acetyl-CoA for lipogenesis [1]. |
| Folate Metabolism | MTHFD1, MTHFD2 | Both | Generates NADPH in both the cytosol and mitochondria, integrated with one-carbon unit metabolism for nucleotide synthesis [7] [1]. |
| Transhydrogenation | PntAB (NADH->NADPH), UdhA (NADPH->NADH) | Cytosol/Membrane | Does not generate de novo NADPH but balances the NADH/NADPH ratio. PntAB is proton-motive force driven [6]. |
The choice depends on the host organism, the subcellular location of your pathway, and the carbon source. For glucose-based growth, engineering the PPP is often most effective. For glutamine-based metabolism, IDH and malic enzyme may be more significant.
Q2: How can I quantitatively measure the NADPH/NADP+ ratio in my engineered cells?
A standard protocol involves rapid sampling and extraction to preserve the in vivo redox state, followed by HPLC-UV analysis [10].
- Sampling & Quenching: Quickly sample cell broth (e.g., 4 mL) directly into a tube containing cold perchloric acid, thoroughly mixing to immediately halt metabolism and stabilize cofactors. At acidic pH, the oxidized forms (NADP+) are stable [10].
- Neutralization: Centrifuge the sample and neutralize the supernatant with appropriate amounts of K~2~HPO~4~ and KOH.
- HPLC Analysis: Analyze the extract using HPLC-UV with a reversed-phase column (e.g., LiChrospher RP-18). A gradient elution with a phosphate-TBAHS buffer system is typically used to separate and quantify the different cofactors [10].
Q3: My product requires NADPH, but I observe redox stress and poor growth. What strategies can help?
This indicates a strong imbalance. Consider:
- Dynamic Regulation: Implement regulatory systems that decouple pathway expression from growth, expressing your NADPH-consuming pathway heavily only after biomass accumulation.
- Synthetic Transhydrogenase Cycles: Design synthetic cycles that use excess ATP or another energy source to drive NADPH formation without being directly coupled to carbon catabolism [9].
- Cofactor Specificity Engineering (CSE): Re-engine your pathway enzymes to accept NADH instead of NADPH, thereby shifting the redox burden to the more abundant NAD(H) pool [9].
Q4: Are there computational tools to predict NADPH flux and guide engineering strategies?
Yes, in silico model-driven approaches are central to modern cofactor engineering.
- Flux Balance Analysis (FBA): Use genome-scale metabolic models (GEMs) to predict intracellular fluxes and identify enzyme targets whose cofactor specificity switching (CSE) will improve overall NADP(H) turnover [9].
- Flux-Sum Analysis: A metabolite-centric approach to understand the metabolic network and identify key nodes for engineering [9].
- Cofactor Modification Analysis (CMA): A computational framework to systematically identify cofactor specificity engineering targets for strain improvement [9].
Experimental Protocol: Enhancing NADPH Supply via the Pentose Phosphate Pathway
Aim: To engineer an E. coli host for increased NADPH supply to support the production of NADPH-demanding compounds (e.g., 5-Methyltetrahydrofolate [8]).
Materials:
- Strain: E. coli BL21(DE3) or other relevant production strain.
- Plasmids: Overexpression vectors for zwf (G6PD) and gnd (PGD).
- Knockout Kit: For deleting pfkA (6-phosphofructokinase 1).
Method:
- Strain Construction:
- Knockout: Delete the pfkA gene from the chromosome to reduce glycolytic flux and increase the pool of glucose-6-phosphate [8].
- Overexpression: Co-transform the strain with plasmids overexpressing zwf and gnd under strong, inducible promoters (e.g., P~Trc~ or P~T7~).
- Cultivation:
- Grow the engineered strain in a defined minimal medium (e.g., M9) with glucose as the sole carbon source.
- Induce gene expression at mid-log phase.
- Validation & Analysis:
- Measure NADPH/NADP+: Use the HPLC protocol described in FAQ A2 to confirm an increased NADPH/NADP+ ratio.
- Quantify Product: Measure the titer of your target product (e.g., via HPLC or LC-MS) to assess the impact of increased NADPH supply.
- Flux Analysis (Optional): Perform ^13^C-metabolic flux analysis (^13^C-MFA) using labeled glucose (e.g., 1-^13^C-glucose) to quantitatively confirm the increased flux through the PPP oxidative phase [10].
The following workflow diagram illustrates this protocol:
Visualization of NADPH Metabolism and Engineering Nodes
The diagram below maps the central roles of NADPH and key engineering targets for balancing its homeostasis in a cellular context.
The Scientist's Toolkit: Key Reagents for NADPH Research
Table 2: Essential Reagents and Strains for NADPH Engineering Experiments
| Research Reagent / Tool | Function / Description | Example Use Case |
|---|---|---|
| Glucose-6-Phosphate Dehydrogenase (G6PD) | Catalyzes the first, rate-limiting step of the PPP, producing NADPH [11]. | Overexpression to boost cytosolic NADPH supply [8]. |
| NAD Kinase (NADK) | Phosphorylates NAD+ to generate NADP+, the precursor for NADPH [7]. | Overexpression to increase the total cellular pool of NADP(H) [7]. |
| Membrane-bound Transhydrogenase (PntAB) | Converts NADH and NADP+ to NAD+ and NADPH, coupling to proton translocation [6]. | Balancing cofactor pools when NADH is abundant but NADPH is limiting [8] [6]. |
| Formate Dehydrogenase (FDH) | Oxidizes formate to CO₂, reducing NAD(P)+ to NAD(P)H [8]. | External regeneration of NAD(P)H using inexpensive sodium formate as an electron donor [8]. |
| ^13^C-Labeled Glucose (e.g., 1-^13^C) | Tracer for metabolic flux analysis (^13^C-MFA). | Quantifying in vivo flux through the PPP versus glycolysis [10]. |
| Engineered E. coli Strains | Hosts with modified central metabolism (e.g., ΔpfkA, ΔldhA, etc.) [8] [10]. | Providing a chassis with optimized carbon flux toward NADPH-generating pathways [8] [10]. |
ATP/ADP as Universal Energy Currency Driving Biosynthetic Reactions
Adenosine Triphosphate (ATP) serves as the universal energy currency in all living cells, providing the fundamental driving force for biosynthetic reactions. In metabolic engineering, managing the balance between ATP and its counterpart ADP, along with redox cofactors like NADPH, is critical for optimizing pathway efficiency in engineered biological systems. This technical support center provides practical guidance for researchers and scientists troubleshooting cofactor balance issues in engineered pathways, offering proven methodologies to diagnose and resolve common experimental challenges.
Frequently Asked Questions (FAQs)
1. What makes ATP the "universal energy currency" rather than other nucleotides like GTP or UTP? ATP is uniquely suited as the primary energy currency due to its molecular structure and thermodynamic properties. The hydrolysis of ATP to ADP releases significant energy (-30.5 kJ/mol or -7.3 kcal/mol) that drives cellular processes [12] [13]. This intermediate energy value positions ATP perfectly to phosphorylate lower-energy compounds while itself being regenerated by higher-energy compounds [14] [13]. Though other nucleotides (GTP, UTP, CTP) participate in specialized metabolic reactions, ATP's ability to readily donate single phosphates, two phosphates, or its adenosine moiety makes it uniquely versatile for energy transfer [15].
2. Why is cofactor balance particularly crucial in engineered metabolic pathways? Engineered pathways disrupt native cellular homeostasis, creating cofactor imbalances that compromise efficiency. Introducing synthetic pathways alters the careful balance of ATP/ADP and NAD(P)/NAD(P)H pools that cells maintain through evolution [16]. Even small changes in these cofactor pools can have wide effects, potentially leading to partial or complete disruption of cellular physiology [16]. Proper cofactor balance ensures that synthetic pathways don't create metabolic bottlenecks that divert resources toward wasteful cycles or biomass formation instead of target compound production [16].
3. How can I troubleshoot inconsistent ATP measurement results in my experiments? Inconsistent ATP measurements often stem from methodological errors rather than biological factors:
- Technique Issues: Apply consistent pressure and use overlapping "Z" pattern when swabbing surfaces [17]
- Timing: Wait 10-15 minutes after cleaning before swabbing, as residual sanitizers interfere with ATP detection [17]
- Sample Integrity: Avoid touching swab tips and ensure proper storage conditions (typically 36–46°F/2–8°C) [17]
- Equipment Maintenance: Regularly clean sensors and calibrate luminometers according to manufacturer specifications [17]
4. What strategies exist for reversing NAD/NADP cofactor specificity in enzymes? A structure-guided, semi-rational strategy called CSR-SALAD (Cofactor Specificity Reversal - Structural Analysis and LibrAry Design) has proven effective for reversing enzymatic nicotinamide cofactor utilization [18]. This approach involves:
- Enzyme structural analysis to identify specificity-determining residues
- Design and screening of focused mutant libraries
- Recovery of catalytic efficiency through compensatory mutations [18] The method successfully inverted cofactor specificity in four structurally diverse NADP-dependent enzymes: glyoxylate reductase, cinnamyl alcohol dehydrogenase, xylose reductase, and iron-containing alcohol dehydrogenase [18].
Troubleshooting Guides
Problem: Low Product Yield in Engineered Pathways
Potential Causes and Solutions:
Table: Cofactor Balance Indicators and Interventions
| Observation | Potential Imbalance | Intervention Strategy |
|---|---|---|
| Reduced product yield with increased biomass | ATP surplus | Engineer ATP-requiring (negative ATP yield) pathways [16] |
| Accumulation of metabolic waste products | Redox imbalance (NAD(P)H) | Modulate PPP flux via gndA or gsdA overexpression [19] |
| Inconsistent performance across conditions | Cofactor specificity mismatch | Implement CSR-SALAD for specificity reversal [18] |
| Stunted growth with pathway activity | ATP/redox deficit | Introduce NADP-dependent GAPDH to generate NADPH instead of NADH [19] |
Diagnostic Protocol:
- Quantify Cofactor Pools: Measure intracellular ATP/ADP and NADPH/NADP+ ratios using standard enzymatic assays
- Flux Analysis: Employ 13C metabolic flux analysis to determine PPP versus EMP pathway utilization [19]
- Computational Modeling: Use Constraint-Based Modelling and co-factor balance assessment (CBA) to predict network-wide effects of pathway engineering [16]
Problem: Enzyme Cofactor Specificity Mismatch
Engineering Workflow:
Implementation Steps:
- Structural Analysis: Input enzyme structure into CSR-SALAD web tool to identify residues contacting the 2' moiety of NAD/NADP [18]
- Library Design: Generate focused mutant libraries using degenerate codons at specificity-determining positions [18]
- Specificity Screening: Screen for activity with non-preferred cofactor using high-throughput methods
- Activity Recovery: Introduce compensatory mutations around adenine ring binding pocket to restore catalytic efficiency [18]
- Pathway Integration: Test engineered enzyme in full pathway context to verify performance
Problem: Inadequate NADPH Supply for Biosynthesis
Enhancement Strategies:
Table: NADPH Generation Enzymes for Cofactor Engineering
| Enzyme | Gene | Pathway | Effect on NADPH | Impact on Production |
|---|---|---|---|---|
| Glucose-6-phosphate dehydrogenase | gsdA | Pentose Phosphate | Moderate increase | Variable (can be negative) [19] |
| 6-phosphogluconate dehydrogenase | gndA | Pentose Phosphate | 45% pool increase | 65% yield increase [19] |
| NADP-dependent malic enzyme | maeA | Reverse TCA cycle | 66% pool increase | 30% yield increase [19] |
| NADP-dependent glyceraldehyde-3-phosphate dehydrogenase | GapN | Glycolysis | Significant (theoretical) | 70-120% yield improvement in C. glutamicum [19] |
Experimental Protocol for NADPH Enhancement:
- Gene Selection: Choose appropriate NADPH-generating enzyme based on host organism and pathway constraints
- Expression Optimization: Use tunable systems (e.g., Tet-on) to control expression levels [19]
- Flux Analysis: Employ chemostat cultures with metabolome analysis to verify NADPH pool increases [19]
- Production Correlation: Measure target compound yield to validate engineering success
Research Reagent Solutions
Table: Essential Materials for Cofactor Balance Research
| Reagent/Kit | Application | Key Features | Considerations |
|---|---|---|---|
| Hygiena UltraSnap ATP swabs | ATP monitoring | Room temperature storage | Check expiration dates; proper storage critical [17] |
| EnSURE Touch luminometer | RLU measurement | Built-in diagnostics | Regular sensor cleaning required [17] |
| CSR-SALAD web tool | Cofactor specificity reversal | Automated structural analysis | Free online resource [18] |
| Genome-scale metabolic models | Cofactor balance prediction | Systems-level analysis | Requires computational expertise [16] |
Advanced Methodologies
Computational Cofactor Balance Assessment
Protocol:
- Model Construction: Use established stoichiometric models (e.g., E. coli core model) [16]
- Pathway Implementation: Introduce heterologous reactions with correct stoichiometry
- Balance Analysis: Apply FBA-based co-factor balance assessment (CBA) algorithm [16]
- Flux Validation: Compare predictions with 13C-MFA data where available
- Futile Cycle Identification: Manually constrain models to eliminate unrealistic cycling [16]
Metabolic Engineering for Cofactor Balancing
Integrated Workflow:
Implementation Notes:
- Strategy Selection: Choose between NADPH generation enhancement (PPP enzymes), cofactor specificity reversal, or ATP balancing based on diagnostic results
- Genetic Implementation: Use CRISPR/Cas9 for precise genome editing and tunable promoters for fine-tuned expression [19]
- Analytical Validation: Combine metabolomics (cofactor pools), fluxomics (pathway utilization), and product quantification for comprehensive assessment
- Iterative Refinement: Apply DBTL (Design-Build-Test-Learn) cycles to progressively improve cofactor balance and pathway performance [19]
Cofactor balance, particularly of ATP and NADPH, is a fundamental requirement for the maintenance of metabolism, energy generation, and growth in engineered biological systems [20]. Metabolic compartmentalization—the spatial and temporal separation of pathways and components—is a key organizational principle that cells use to manage this balance [21]. For metabolic engineers, understanding and engineering this compartmentalization is essential for designing efficient microbial cell factories, as it fulfills three primary functions or "pillars": establishing unique chemical environments for reactions, protecting the cell from reactive intermediates, and providing precise regulatory control over metabolic pathways [21]. This technical support center provides troubleshooting guides, experimental protocols, and key resources to help researchers navigate the challenges of managing cofactor pools in engineered pathways.
Troubleshooting FAQs
1. My pathway has the necessary enzymes expressed, but product titers are low and growth is impaired. Could cofactor imbalance be the issue?
- Answer: Yes, this is a classic symptom of cofactor imbalance. Your pathway may be consuming NADPH or ATP faster than native metabolism can regenerate it, or vice-versa.
- Solutions:
- Quantify the Demand: First, calculate the theoretical ATP:NADPH demand of your engineered pathway. A recent meta-analysis revealed that pathways like starch and sucrose synthesis make a notable contribution to the overall energy demand, which can counterbalance the demand from other routes like photorespiration [22].
- Engineer Cofactor Regeneration: Implement synthetic cofactor regeneration cycles. For NADPH supply, consider overexpressing genes from the oxidative pentose phosphate pathway (e.g., G6PDH, 6PGDH) or introducing a transhydrogenase [23].
- Modify Cofactor Preference: Use protein engineering to alter the cofactor specificity of a key enzyme in your pathway from NADPH to NADH, as the latter is often more abundant under anaerobic conditions [23] [20].
2. How can I determine if subcellular localization is causing a bottleneck in my cofactor-dependent pathway?
- Answer: A compartmentalization bottleneck is likely if an enzyme shows high activity in vitro but not in vivo, or if metabolic flux models predict higher yields than are experimentally achieved.
- Solutions:
- Re-target Enzymes: Re-locate your pathway enzymes to a cellular compartment with a more favorable cofactor pool. For instance, engineering the functional expression of prokaryotic P450 enzymes in yeast mitochondria, rather than the cytosol, significantly improved the production of 10-hydroxy-2-decenoic acid by leveraging the mitochondrial environment and NADPH supply [24].
- Create Synthetic Compartments: Use synthetic biology tools to create engineered metabolic organelles or protein scaffolds to concentrate pathway enzymes and cofactors, thus enhancing flux and preventing cross-talk with native metabolism [21].
3. I have engineered a cofactor regeneration system, but it has caused unexpected reductions in the synthesis of my target product. Why?
- Answer: Cofactor perturbations can have widespread, systemic effects because redox cofactors are among the most highly connected metabolites in the network [20]. Altering one pool can divert key precursors away from your target pathway.
- Solutions:
- Analyze Precursor Availability: Check the levels of key pathway precursors like acetyl-CoA and α-keto acids. A study in Saccharomyces cerevisiae showed that redox imbalance modified the formation of all volatile compounds from the same biochemical pathway due to its impact on central carbon metabolism intermediates [20].
- Perturbation Analysis: Use a dedicated biological tool, like the overexpression of a native NADH-dependent or engineered NADPH-dependent 2,3-butanediol dehydrogenase in the presence of acetoin, to specifically perturb cofactor balance and analyze the resulting metabolic changes without permanently altering central metabolism [20].
4. My microbial host shows poor energy efficiency after pathway introduction. Which reactions are the most energy-expensive?
- Answer: Energy failure may not be a general deficiency in producing ATP, but a failure to recoup the ATP cost of certain processes. A genome-wide CRISPR screen identified hexokinase 2 (HK2), the first enzyme in glycolysis, as one of the greatest ATP consumers in a human cell line [25].
- Solutions:
- Identify Key Consumers: Use computational models or literature mining to identify the most energy-expensive steps in your host's metabolic network.
- Modulate Consumers: Consider down-regulating high-consumption reactions that are non-essential for your production objective. The screen revealed that suppressing HK2 or its binding partner VDAC1 increased ATP levels under respiratory conditions [25].
Experimental Protocols
Protocol 1: Quantifying In Vivo Cofactor Pools and Ratios
This protocol describes a method for measuring the absolute concentrations of NAD+, NADH, NADP+, and NADPH in microbial cultures, adapted from a study on redox cofactor perturbations [20].
1. Principle: Rapid quenching of metabolism to preserve in vivo state, followed by metabolite extraction and enzymatic assay or LC-MS/MS quantification.
2. Reagents:
- Quenching Solution: Cold methanol buffer (60% methanol, 70 mM HEPES, pH 7.5) at -40 °C.
- Extraction Buffer: 100% methanol or acetonitrile-based buffer.
- Analytical Standards: Stable isotope-labeled internal standards for NAD+, NADH, NADP+, NADPH.
3. Procedure:
- Step 1: Culture Sampling. Rapidly withdraw a known volume of culture (e.g., 5-10 mL) and immediately mix it with a larger volume of pre-chilled Quenching Solution (-40 °C) to instantaneously halt metabolic activity.
- Step 2: Metabolite Extraction.
- Centrifuge the quenched cells at high speed (e.g., 10,000 x g, 5 min, -20 °C).
- Discard the supernatant and resuspend the cell pellet in cold Extraction Buffer.
- Vortex or sonicate the suspension to lyse cells and release intracellular metabolites.
- Centrifuge again to remove cell debris. The supernatant containing the metabolites is used for analysis.
- Step 3: Analysis.
- Option A (Enzymatic): Use specific cycling assays for each cofactor. For example, quantify NADPH using glutathione reductase and 5,5'-dithio-bis-(2-nitrobenzoic acid) (DTNB).
- Option B (LC-MS/MS): This is the preferred method for simultaneous and specific quantification. Separate the extracted cofactors using hydrophilic interaction liquid chromatography (HILIC) and detect them via tandem mass spectrometry. Use the internal standards for precise quantification.
4. Data Analysis: Calculate the concentration of each cofactor (in nmol/gDCW) and determine the ratios NAD+/NADH and NADP+/NADPH. These ratios are key indicators of the cellular redox state.
Protocol 2: Engineering Compartmentalization for Cofactor-Dependent Pathways
This protocol outlines the strategy used to successfully produce 10-HDA in S. cerevisiae by harnessing mitochondrial compartmentalization [24].
1. Principle: Re-locate a cofactor-intensive pathway to an organelle with a favorable environment to enhance flux, stability, and cofactor availability.
2. Reagents:
- Expression Vectors: Plasmids with promoters for strong, constitutive expression (e.g., TEF1, PGK1).
- Targeting Sequences: DNA sequences encoding mitochondrial targeting signals (MTS) for N-terminal fusion to enzymes.
- Chaperone Plasmids: Vectors for co-expressing chaperone proteins (e.g., mitochondrial Hsp60) to assist in the folding of recombinant enzymes [24].
3. Procedure:
- Step 1: Pathway Design and Gene Synthesis. Design the heterologous pathway (e.g., the β-oxidation pathway for 10-HDA). Codon-optimize genes for your host.
- Step 2: Addition of Targeting Sequences. Fuse mitochondrial targeting signals to the N-terminus of the enzymes you wish to localize to the mitochondria. For the P450 enzyme in the 10-HDA study, this was critical for functional expression [24].
- Step 3: Chassis Engineering.
- Transform the host with the engineered gene constructs.
- To enhance the mitochondrial NADPH supply, consider overexpressing enzymes that generate NADPH in the mitochondria, such as a mitochondrial-targeted NADP+-dependent isocitrate dehydrogenase.
- Step 4: Fed-Batch Fermentation.
- To mitigate substrate toxicity, use a fed-batch strategy with a less toxic substrate precursor. In the 10-HDA example, using ethyl decanoate instead of decanoic acid increased production 7.5-fold [24].
- Monitor product titer and cell density throughout the fermentation.
The Scientist's Toolkit: Research Reagent Solutions
Table 1: Essential reagents and tools for engineering cofactor systems and compartmentalization.
| Reagent/Tool | Function & Application | Key Consideration |
|---|---|---|
| CRISPRi/a System [25] | Genome-wide screening to identify genes that regulate ATP levels (the "ATPome") or other cofactors. | Enables discovery of both ATP consumers (CRISPRi) and genes that boost ATP when overexpressed (CRISPRa). |
| Isotopically Non-Stationary Metabolic Flux Analysis (INST-MFA) [22] | Quantitative mapping of in vivo metabolic reaction rates (fluxes) in central carbon metabolism. | Essential for experimentally measuring the energy (ATP:NADPH) demand of different pathways under various conditions. |
| 2,3-Butanediol Dehydrogenase (Bdh) [20] | A dedicated biological tool for targeted perturbation of NADH or NADPH balance. An engineered NADPH-dependent version allows for specific cofactor manipulation. | Used with acetoin to create a controlled sink for reduced cofactors, allowing study of the metabolic response. |
| Mitochondrial Targeting Signal (MTS) [24] | Peptide sequence fused to enzymes to re-target them from the cytosol to the mitochondrial matrix. | Used to leverage unique chemical environments and potentially higher cofactor concentrations in organelles. |
| Genome-Scale Metabolic Models (GEMs) [26] | Computational stoichiometric models of metabolism used for in silico prediction of metabolic fluxes. | Use with algorithms like OptORF to predict gene knockouts that optimize cofactor balance and product yield. |
Visualizing Key Concepts and Workflows
Pillars of Compartmentalization
Mitochondrial Engineering Workflow
Cofactor Engineering Strategies and Data
Table 2: Summary of cofactor engineering approaches and their outcomes.
| Engineering Strategy | Specific Action | Organism | Key Outcome / Impact |
|---|---|---|---|
| Self-Balance [23] | Modifying overflow metabolism (e.g., glycerol production) | S. cerevisiae | Automatically maintains redox balance by rerouting central carbon metabolism. |
| Substrate Balance [23] | Providing electron acceptors or altering culture conditions (e.g., oxygen) | Microbes | Achieves optimal NADH/NAD+ ratio by modifying NADH reoxidation. |
| Synthetic Balance [23] | Protein engineering to switch cofactor preference (NADPHNADH) | Various | Rebalances oxidoreduction potential in imbalanced pathways, improving product yield. |
| Compartmentalization [24] | Rewiring β-oxidation + P450 expression in mitochondria | S. cerevisiae | Achieved 298.6 mg/L of 10-HDA, the highest titer in yeast, by leveraging compartmentalization. |
| CRISPR-based Screening [25] | Inhibition (CRISPRi) of hexokinase 2 (HK2) | Human K562 cells | Increased ATP levels under respiratory conditions by suppressing a major ATP consumer. |
The Critical Link Between Cofactor Imbalance and Reduced Bioproduction Yields
Troubleshooting Guides
Guide 1: Diagnosing and Resolving NADPH Limitation
Problem: Low product yield in pathways dependent on NADPH, such as fatty acid or amino acid biosynthesis.
Question: How can I determine if my production strain is experiencing NADPH limitation, and what are the primary strategies to overcome it?
Answer: NADPH limitation is a common bottleneck in reductive biosynthetic pathways. Diagnosis can involve checking for an accumulation of pathway intermediates or using genetically encoded biosensors to monitor the intracellular NADPH/NADP+ ratio [27]. The solutions below are ranked from the most common and straightforward to the more advanced.
1. Reinforce the Pentose Phosphate Pathway (PPP): This is the primary source of cytosolic NADPH. * Action: Overexpress the genes zwf (glucose-6-phosphate dehydrogenase) and gnd (6-phosphogluconate dehydrogenase) [27]. * Example: In E. coli, enhancing the PPP flux has been used to improve the production of products like poly-3-hydroxybutyrate (PHB) [27].
2. Employ a Cofactor Boosting System: A versatile approach to generally enhance the pool of cofactor precursors. * Action: Implement a system like xylose reductase with lactose (XR/lactose). This system increases sugar phosphate pools, which are connected to the biosynthesis of NADPH, FAD, FMN, and ATP [28]. * Example: The XR/lactose system increased productivities in fatty alcohol biosynthesis, bioluminescence generation, and alkane biosynthesis by 2-4 fold in E. coli [28].
3. Engineer Cofactor Specificity: * Action: Replace a native NADH-dependent enzyme in your pathway with a non-native NADPH-dependent version. * Example: Replacing the native E. coli glyceraldehyde-3-phosphate dehydrogenase (GAPD, gapA) with the NADPH-dependent GAPD from Clostridium acetobutylicum (gapC) can increase NADPH availability for product synthesis [29].
4. Implement Dynamic Regulation: * Action: Use genetically encoded biosensors to dynamically regulate the NADPH/NADP+ balance in real-time, moving beyond static overexpression [27]. * Example: The SoxR biosensor in E. coli or the NERNST biosensor can be used to monitor and respond to the intracellular NADPH/NADP+ status [27].
Guide 2: Addressing NADH/NAD+ Imbalance
Problem: Reduced cell growth, metabolic arrest, or byproduct accumulation due to an overload of NADH, often a problem in cyanobacteria or pathways producing excess reducing equivalents.
Question: My pathway generates excess NADH, leading to reductive stress and poor performance. How can I rebalance the NADH/NAD+ pool?
Answer: An excess of NADH can inhibit critical metabolic enzymes and waste energy. The goal is to increase NADH oxidation or reduce its net production.
1. Introduce NADH Oxidase (Nox): * Action: Express a heterologous NADH oxidase to convert NADH to NAD+. * Example: Expression of SpNox from Streptococcus pyogenes in E. coli created an NAD+ regeneration system that helped achieve a high pyridoxine titer of 676 mg/L by alleviating NADH surplus [30] [31].
2. Swap Cofactor Specificity in Glycolysis: * Action: Modify central metabolism to reduce NADH generation. Substitute the native NADH-producing glyceraldehyde-3-phosphate dehydrogenase with a NADPH-producing version. * Example: In E. coli, this swap reduces glycolytic NADH production, helping to balance cofactors for targets like pyridoxine [30].
3. Leverage Native Pathways in Specialized Organisms: * Action: In cyanobacteria, which have an inherently NADPH-rich pool, express NADPH-dependent versions of enzymes that are normally NADH-dependent. * Example: Changing the cofactor specificity of enzymes in cyanobacteria from NADH to NADPH can overcome the innate cofactor imbalance and enhance production of chemicals like ethanol or lactic acid [32].
Guide 3: Overcoming ATP and Energy Cofactor Deficits
Problem: Stalled biosynthesis in ATP-intensive pathways, such as luciferase-based systems or the production of certain polymers.
Question: How can I enhance the ATP supply to drive energy-intensive bioproduction?
Answer: 1. Utilize a Generic Cofactor Booster: * Action: Systems that enhance sugar phosphate pools, like the XR/lactose system, also propagate benefits to ATP biosynthesis [28]. * Example: The XR/lactose system enhanced bioluminescence light generation in E. coli, a process with high demand for ATP [28].
2. Engineer ATP Regeneration Pathways: * Action: Overexpress enzymes like polyphosphate kinase to regenerate ATP from ADP and polyphosphate [28].
Table 1: Summary of Common Cofactor Imbalances and Solutions
| Cofactor Issue | Key Symptoms | Recommended Engineering Strategies |
|---|---|---|
| NADPH Limitation | Low yield of reduced products (e.g., alcohols, fatty acids); Accumulation of oxidized precursors. | • Reinforce Pentose Phosphate Pathway (zwf, gnd overexpression) [27]• Use XR/lactose boosting system [28]• Implement Cofactor Swapping (e.g., gapC) [29] |
| NADH/NAD+ Imbalance | Reductive stress; Impaired cell growth; Byproduct formation (e.g., lactate, ethanol). | • Express NADH Oxidase (Nox) [30] [31]• Reduce NADH production via glycolytic enzyme swaps [30]• Use NADPH-dependent enzymes in cyanobacteria [32] |
| ATP Deficit | Stalled anabolic processes; Low yields in energy-intensive pathways (e.g., luminescence). | • Deploy XR/lactose system [28]• Engineer ATP regeneration (e.g., polyphosphate kinase) [28] |
Frequently Asked Questions (FAQs)
FAQ 1: What is the fundamental difference between NADH and NADPH in cellular metabolism, and why does it matter for metabolic engineering?
While both are reducing equivalents, they are functionally segregated. NADH is primarily involved in catabolic reactions to generate ATP, whereas NADPH is primarily used for anabolic (biosynthetic) reactions and antioxidant defense [1]. This division allows the cell to independently manage energy production and biomass synthesis. Imbalances occur when engineered pathways disrupt this natural partition, for example, by consuming too much NADPH and starving native biosynthesis, or by generating excess NADH that cannot be re-oxidized [23] [29].
FAQ 2: Beyond the PPP, what are other major sources of NADPH in the cell that I can engineer?
Two other crucial sources are:
- Isocitrate Dehydrogenase (IDH): Both the cytosolic (IDH1) and mitochondrial (IDH2) enzymes generate NADPH from isocitrate in the TCA cycle [1] [27].
- Malic Enzyme (ME): The cytosolic (ME1) and mitochondrial (ME3) enzymes generate NADPH by converting malate to pyruvate [1]. Engineering these pathways, such as by overexpressing IDH genes from other species, has been shown to enhance NADPH supply [27].
FAQ 3: What are the pros and cons of static vs. dynamic regulation for cofactor balancing?
- Static Regulation (e.g., gene knockout, constitutive overexpression): This is simpler to implement but cannot respond to changing cellular conditions. It often leads to metabolic burdens or new imbalances because the regulation is fixed [27].
- Dynamic Regulation: This uses biosensors to monitor cofactor levels (like the NADPH/NADP+ ratio) and adjust gene expression in real-time. It is more sophisticated and maintains metabolic flexibility, preventing imbalance by responding to the cell's actual needs [27]. This is considered a more advanced and robust strategy.
Experimental Protocols
Protocol 1: Implementing the XR/Lactose Cofactor Boosting System
Objective: To enhance the intracellular pool of multiple cofactors (NADPH, FAD, FMN, ATP) in E. coli to support engineered pathways.
Background: This system uses xylose reductase (XR) to reduce the hydrolyzed products of lactose (glucose and galactose) into sugar alcohols. Their metabolism leads to the accumulation of sugar phosphates, which are precursors for cofactor biosynthesis [28].
Materials:
- Strain: Engineered E. coli BL21(DE3) with your production pathway.
- Plasmid: Vector containing the XR gene from Hypocrea jecorina.
- Inducer: Lactose.
- Culture Media: Standard LB or defined minimal media.
Method:
- Strain Construction: Transform your production host with the plasmid carrying the XR gene.
- Culture Induction:
- Inoculate main culture and grow to mid-log phase.
- Induce protein expression by adding lactose (typical range 2–20 g/L) [28].
- Continue cultivation for a set period (e.g., 6 hours).
- Bioconversion Assay:
- Harvest cells by centrifugation.
- Resuspend the cell pellet in a reaction buffer containing a high concentration of lactose (or other sugar carbon source) to serve as the substrate for the cofactor boosting system.
- Incubate to allow for product formation.
- Analysis: Measure product titer (e.g., fatty alcohols, alkane) using GC-MS or other appropriate methods and compare with a control strain without XR.
Expected Outcome: The XR/lactose system has been shown to increase productivities by 2-4 fold in various systems [28].
Protocol 2: Enhancing NADPH Supply via Pentose Phosphate Pathway Overexpression
Objective: To increase NADPH availability by overexpressing key enzymes in the oxidative PPP.
Background: The enzymes glucose-6-phosphate dehydrogenase (Zwf) and 6-phosphogluconate dehydrogenase (Gnd) are the primary generators of NADPH in the PPP.
Materials:
- Strain: Your production E. coli strain.
- Plasmids: Vectors for expressing zwf and gnd under an inducible promoter.
Method:
- Strain Engineering: Construct a strain overexpressing zwf and gnd. This can be done via chromosomal integration or using plasmids.
- Cultivation: Grow the engineered strain and control in parallel under production conditions.
- Validation and Production:
- Metabolomic Analysis: Use LC-MS to measure levels of PPP intermediates (e.g., glucose-6-phosphate, 6-phosphogluconate) to confirm increased flux.
- Product Measurement: Quantify the target product to assess yield improvement.
Expected Outcome: This strategy has successfully enhanced production of compounds like mevalonate and terpenes by ensuring an ample NADPH supply [27].
Data Presentation
Table 2: Quantitative Impact of Cofactor Engineering on Bioproduction Yields
| Target Product | Host Organism | Cofactor Challenge | Engineering Strategy | Resulting Yield Improvement | Citation |
|---|---|---|---|---|---|
| Fatty Alcohols | E. coli | NADPH, Acetyl-CoA demand | XR/Lactose Cofactor Boosting System | ~3-fold increase (from 58.1 to 165.3 μmol/L/h) | [28] |
| Pyridoxine (Vitamin B6) | E. coli | NADH/NAD+ imbalance | Multiple strategies: NADH oxidase (SpNox), glycolytic enzyme swaps, protein engineering | Titer reached 676 mg/L in shake flask | [30] [31] |
| Theoretical Yields (Various) | E. coli & S. cerevisiae | NADPH limitation | Computational identification of optimal cofactor swaps (e.g., GAPD) | Increased theoretical maximum yield for many native and non-native products | [29] |
| Alkanes / Bioluminescence | E. coli | FAD, FMN, ATP demand | XR/Lactose Cofactor Boosting System | 2-4 fold increase in productivity | [28] |
Pathway and Workflow Visualizations
XR Lactose Cofactor Boosting System
Bioproduction Yield Troubleshooting Guide
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Key Reagents for Cofactor Engineering
| Reagent / Tool | Function / Application | Example Use Case |
|---|---|---|
| Xylose Reductase (XR) | Reduces various sugars (glucose, galactose) to sugar alcohols, feeding into pathways that boost sugar phosphate and cofactor pools. | Core component of the versatile XR/lactose cofactor boosting system [28]. |
| NADH Oxidase (Nox) | Oxidizes NADH to NAD+, regenerating the oxidized cofactor and resolving reductive stress. | SpNox from S. pyogenes used to improve pyridoxine production in E. coli [30] [31]. |
| Cofactor Biosensors | Genetically encoded tools for real-time monitoring of intracellular cofactor ratios (e.g., NADPH/NADP+). | SoxR biosensor in E. coli or the NERNST ratiometric biosensor for dynamic regulation [27]. |
| Non-native GAPD Enzymes | Swaps cofactor specificity in glycolysis. gapC provides NADPH; gapN can reduce NADH production. | gapC from C. acetobutylicum expressed in E. coli to increase NADPH supply [29] [30]. |
| CRISPR-Cas9 System | Enables precise genome editing for gene knockouts, knock-ins, and promoter replacements. | Used for traceless genome editing in E. coli to delete competing genes or integrate pathway genes [31]. |
Engineering Solutions: Strategic Approaches for NADPH and ATP Balance Control
Troubleshooting Guides
FAQ 1: My FBA predictions show high product yields, but my experimental results reveal significant metabolite waste (e.g., acetate). What is the cause and how can I resolve it?
Answer: This discrepancy often arises from cofactor imbalance, where an excess of ATP or NAD(P)H generated by your engineered pathway is dissipated via native metabolic reactions, leading to by-product formation and reduced product yield [33].
- Root Cause: The FBA solution space is underdetermined, allowing for futile cofactor cycles with high flux. These are thermodynamically inefficient cycles where, for example, ATP is hydrolyzed without productive work, or cofactors are cycled to balance redox without contributing to growth or production [33].
- Solution:
- Implement a Co-factor Balance Assessment (CBA) Protocol: Systematically categorize all reactions producing or consuming ATP and NAD(P)H in your model. This helps identify if surplus cofactors are being diverted to by-product formation or maintenance instead of your target product [33] [34].
- Apply Manual Constraints: Based on experimental data (e.g., from 13C-MFA), manually constrain the fluxes of reactions identified as major contributors to futile cycling [33].
- Re-formulate the Objective Function: If biomass formation is the objective, the model may over-prioritize growth by using futile cycles to dump excess energy. Try setting the biosynthetic production as the primary objective to find solutions that minimize futile cycling [33].
FAQ 2: How can I account for uncertainty in biomass composition in my FBA model to improve cofactor balance predictions?
Answer: Uncertainty in the stoichiometric coefficients of the biomass reaction can propagate and affect the accuracy of flux predictions, including those related to cofactor balance [35].
- Root Cause: The biomass reaction is a major sink for ATP and reducing power. Incorrect coefficients can lead to misrepresentation of the energy and redox demands of growth, causing erroneous predictions of product yield and by-product formation [35].
- Solution:
- Apply Conditional Parameter Sampling: When assessing parameter uncertainty, always sample biomass coefficients in a way that maintains the total molecular weight of the biomass composition at 1 g mmol⁻¹. This ensures thermodynamic consistency [35].
- Enforce Elemental and Metabolite Pool Conservation: Impose constraints that conserve elements and metabolite pools, especially when modeling conditions that deviate from steady-state, to prevent unrealistic flux distributions [35].
FAQ 3: What strategies can I use to re-balance NADPH/NADP+ and ATP/ADP pools in an engineered pathway identified as imbalanced by CBA?
Answer: Computational identification of imbalance should be followed by biological re-engineering.
- Root Cause: A synthetic pathway may consume or produce cofactors in a ratio that is incompatible with the host's native metabolism, creating a redox or energy bottleneck [33] [23].
- Solution:
- Promoter Engineering: Fine-tune the expression of cofactor-dependent genes in your pathway to optimize flux and reduce imbalance [23].
- Protein Engineering: Modify enzyme specificity to switch cofactor preference (e.g., from NADH to NADPH) [23].
- Heterologous Enzyme Expression: Introduce alternative, better-balanced pathway variants or enzymes such as transhydrogenases or NADH kinases to regenerate the required cofactors [33] [23].
- Use Genetically Encoded Biosensors: Implement biosensors for ATP/ADP and NAD(P)H/NAD(P)+ to monitor intracellular cofactor levels in real-time and guide your engineering efforts [36].
Experimental Protocols
Protocol 1: Implementing a Co-factor Balance Assessment (CBA) Algorithm
This protocol details the steps to perform a CBA using a core E. coli stoichiometric model, as described in the primary literature [33].
1. Model Modification
- Objective: Introduce the reactions for your synthetic pathway into the host's metabolic model.
- Methodology:
- Use a well-curated model (e.g., the E. coli Core Model).
- Add necessary metabolites and reactions to enable the production of your target compound (e.g., butanol). Define an appropriate objective function, such as maximizing the flux through a target product sink reaction [33].
2. Flux Calculation
- Objective: Obtain a flux distribution for the network.
- Methodology:
- Perform Flux Balance Analysis (FBA) or parsimonious FBA (pFBA) to simulate growth or production under defined conditions.
- Record the flux value (
v) for every reaction in the network [33].
3. CBA Flux Categorization
- Objective: Quantify the contribution of different metabolic processes to the cofactor pools.
- Methodology:
- For each cofactor (e.g., ATP, NADH, NADPH), identify all reactions that involve it.
- Categorize the fluxes of these reactions into five core categories based on their function [33] [34]:
- Cofactor Production: Reactions that generate the cofactor (e.g., ATP in glycolysis).
- Biomass Production: Cofactor consumed for growth.
- Target Production: Net cofactor flux from the introduced synthetic pathway.
- Waste Release: Cofactor flux associated with by-product secretion (e.g., acetate, lactate) or known "futile" cycles.
- Cellular Maintenance: Residual cofactor consumption not accounted for by the above.
The workflow for this protocol is standardized as follows:
Protocol 2: Minimizing Futile Cycles in FBA Predictions
This protocol outlines methods to reduce unrealistic futile cycling identified by the CBA [33].
1. Problem Identification
- Run the CBA protocol. A high flux in the "Waste Release" category, particularly through non-essential ATP-hydrolyzing or NAD(P)H-consuming reactions, indicates probable futile cycling [33] [34].
2. Applying Flux Constraints
- Objective: Manually constrain the model to eliminate unrealistic cycles.
- Methodology:
- Use experimental data from 13C-Metabolic Flux Analysis (MFA) to define physiologically plausible lower and upper bounds for specific reaction fluxes.
- Apply these constraints to your model and re-run the FBA and CBA [33].
3. Alternative Computational Approaches
- Objective: Use built-in algorithms to suppress loop formation.
- Methodology:
- Employ "loopless FBA" or related techniques that explicitly prohibit thermodynamically infeasible cyclic fluxes [33].
- Compare the results with the manually constrained model to identify a robust solution.
The logical process for troubleshooting futile cycles is as follows:
Data Presentation
Table 1: Cofactor Demand and Theoretical Yield of Butanol Production Pathways in E. coli
This table, adapted from a case study, summarizes how different pathway designs affect cofactor balance and theoretical yield [33].
| Model Name | Key Pathway Enzymes | Target Product | ATP Balance (Net) | NAD(P)H Balance (Net) | Relative Yield Potential |
|---|---|---|---|---|---|
| BuOH-0 | AtoB + CP + AdhE2 | Butanol | 0 | -4 | Medium |
| BuOH-1 | NphT7 + CP + AdhE2 | Butanol | -1 | -2 | Higher |
| tpcBuOH | Pathway from M. extorquens | Butanol | 0 | -2 | High |
| BuOH-2 | NphT7 + Ter + AdhE2 | Butanol | -1 | -4 | Lower |
| CROT | Crotonase | Crotonyl-CoA | -1 | 1 | Varies |
| BUTAL | Butyraldehyde dehydrogenase | Butyraldehyde | 0 | -2 | Varies |
Table 2: Research Reagent Solutions for Cofactor Balance Studies
This table lists key reagents and computational tools essential for research in this field.
| Reagent / Tool | Function / Application | Key Feature |
|---|---|---|
| Genetically Encoded ATP/ADP Biosensor | Real-time monitoring of intracellular ATP/ADP ratios in live cells [36]. | Enables dynamic tracking of energy status during production. |
| Genetically Encoded NAD(P)H Biosensor | Real-time monitoring of intracellular NAD(P)H/NAD(P)+ ratios [36]. | Reveals redox challenges in engineered pathways. |
| E. coli Core Model | A simplified stoichiometric model of E. coli metabolism [33]. | Standard platform for implementing FBA and CBA algorithms. |
| Loopless FBA Algorithm | A variant of FBA that eliminates thermodynamically infeasible cyclic fluxes [33]. | Reduces prediction of unrealistic futile cycles. |
| 13C-Metabolic Flux Analysis (13C-MFA) | Experimental technique to measure intracellular metabolic fluxes [33]. | Provides data for constraining and validating FBA models. |
Protein Engineering for Cofactor Specificity Switching and Optimization
Frequently Asked Questions (FAQs)
FAQ 1: Why is switching an enzyme's cofactor specificity from NADPH to NADH often desirable in metabolic engineering?
The primary motivation is cost and stability. NADH is generally less expensive and more stable than NADPH, making processes that rely on it more economical for large-scale applications like chemical and pharmaceutical manufacturing [37]. Furthermore, natural metabolic pathways in production hosts like E. coli or yeast often generate different ratios of these cofactors. Switching specificity allows you to balance cofactor pools within the cell, preventing a build-up of one cofactor and a shortage of another, which can enhance the flux through your engineered pathway and increase product yield [38] [39].
FAQ 2: When I successfully change my enzyme's cofactor preference, why does the catalytic efficiency often decrease, and how can I mitigate this?
A loss in catalytic efficiency is a common challenge because mutations in the cofactor-binding pocket can disrupt optimal geometry and interactions. You can mitigate this by using advanced protein engineering strategies:
- Loop Exchange: Replace entire loops in the cofactor-binding site rather than making single point mutations. This approach has generated some of the best results in terms of both specificity reversal and retained efficiency [38].
- Computational Design: Use algorithms to predict mutations that introduce favorable interactions with the new cofactor (e.g., introducing basic residues to bind NADPH's phosphate group) while minimizing structural disruption [38].
- Directed Evolution: After making initial rational mutations to switch specificity, perform iterative rounds of mutagenesis and screening to select for variants that have regained high catalytic activity [37].
FAQ 3: What are the major sources of NADPH in a typical microbial cell that I can engineer to improve supply?
You can target several key pathways to enhance NADPH regeneration [1]:
- Pentose Phosphate Pathway (PPP): The oxidative phase is a major source. Overexpression of glucose-6-phosphate dehydrogenase (G6PDH) is a common strategy.
- TCA Cycle-related Enzymes: Cytosolic or mitochondrial isoforms of isocitrate dehydrogenase (IDH1/IDH2) and malic enzyme (ME1/ME3) can be engineered for higher flux.
- Transhydrogenation: The membrane-bound transhydrogenase (PntAB) can be manipulated to shift the balance between NADH and NADPH.
- One-Carbon Metabolism: This pathway also contributes to NADPH generation in both the cytosol and mitochondria.
FAQ 4: Beyond switching cofactors, what other cofactor engineering strategies can I use?
A powerful alternative is "Cofactor Engineering of a Network's Cofactor Preference." Instead of re-engineering a single enzyme, you identify and replace multiple enzymes within your heterologous pathway with isofunctional enzymes that naturally possess the desired cofactor specificity. For example, to change a pathway from NADH- to NADPH-dependency, you can replace NADH-dependent enzymes with homologs that use NADPH, thereby creating a pathway that aligns with the host's native cofactor supply [37].
Troubleshooting Guides
Troubleshooting Guide 1: Cofactor Specificity Reversal
| Problem | Possible Cause | Solution |
|---|---|---|
| No activity with new cofactor | Mutations completely disrupted cofactor binding pocket. | Verify binding pocket structure; try a different mutagenesis strategy like loop exchange [38]. |
| Severe loss of catalytic efficiency | Mutations suboptimally positioned, affecting transition state. | Employ directed evolution or computational design to improve efficiency after initial switch [38]. |
| Incomplete specificity switch | Mutations insufficient to overcome wild-type preference. | Introduce additional targeted mutations; analyze successful case studies for your enzyme class [38]. |
| Poor protein expression or stability | Mutations caused misfolding or aggregation. | Include protein stability predictions in design; use lower expression temperatures or chaperones. |
Troubleshooting Guide 2: Managing Cofactor Balance in Engineered Pathways
| Problem | Possible Cause | Solution |
|---|---|---|
| Low product titer despite high pathway enzyme expression | Cofactor imbalance (e.g., NADPH depletion). | Use genomic tools like CRISPRi to downregulate native NADPH-consuming genes [4]. |
| Accumulation of metabolic intermediates | Cofactor imbalance halting pathway flux. | Overexpress key enzymes from PPP (e.g., G6PDH) to boost NADPH supply [1]. |
| Reduced cell growth or viability | Engineering caused ATP/ADP or NADPH/NADP+ imbalance. | Use dynamic regulation systems (e.g., quorum-sensing) to downregulate ATP-consuming processes only at high cell density [4]. |
| Inability to monitor cofactor levels in vivo | Lack of real-time, non-destructive monitoring. | Employ genetically encoded biosensors for ATP/ADP or NADPH/NADP+ to track cofactor dynamics in live cells [36]. |
Experimental Protocols
Protocol 1: A Rational Design Workflow for Switching Cofactor Specificity from NADPH to NADH
Methodology: This protocol outlines a standard rational design approach for cofactor engineering, based on a review of over 100 enzyme engineering studies [38].
Materials:
- Template: Wild-type enzyme structure (X-ray crystal structure preferred)
- Software: Molecular visualization software (e.g., PyMOL), computational protein design software (e.g., Rosetta)
- Strains: E. coli BL21(DE3) or similar for protein expression
- Reagents: Cofactors (NADPH, NADH), substrates, site-directed mutagenesis kit, chromatography media for protein purification
Procedure:
- Structural Analysis: Identify the cofactor-binding pocket in your enzyme structure, typically a Rossman fold. Note key residues that interact with the 2'-phosphate group of NADPH [38].
- Target Identification: pinpoint acidic (negatively charged) residues that form hydrogen bonds with the 2'- and 3'-hydroxyl groups of NADPH's adenine ribose. Also, look for residues that create a positively charged pocket to accommodate the phosphate [38].
- Mutagenesis Design:
- The general strategy is to remove acidic residues and incorporate basic residues.
- Common mutations involve substituting Asp or Glu for Ala, Ser, or Gly to eliminate repulsion against NADH.
- In some cases, introducing Arg or Lys can help form a hydrogen bond with the 2'-phosphate of NADPH if switching to NADPH preference [38].
- Generate Variants: Use site-directed mutagenesis to create the designed mutants.
- Expression & Purification: Express the mutant proteins in your host and purify them using affinity chromatography.
- Kinetic Characterization: Determine the kinetic parameters (
kcatandKm) for both the wild-type and mutant enzymes with both NADPH and NADH. Calculate the Coenzyme Specificity Ratio and Relative Catalytic Efficiency to quantify your success [38].
Protocol 2: CRISPRi Screening for Identifying Cofactor-Consuming Gene Targets (CECRiS)
Methodology: This protocol describes the Cofactor Engineering based on CRISPRi Screening (CECRiS) strategy used to identify native NADPH- and ATP-consuming genes whose repression improves product formation [4].
Materials:
- Strain: Production host (e.g., E. coli 4HPAA-2 from the study) containing your biosynthetic pathway.
- Plasmids: dCas9* plasmid and sgRNA-expression plasmids targeting NADPH-consuming or ATP-consuming genes.
- Media: Selective LB broth or defined production medium.
- Equipment: Shake flasks, bioreactor, HPLC or GC for product quantification.
Procedure:
- sgRNA Library Design: Design sgRNAs to bind the non-template strand ~100 bp downstream of the start codon of all target genes (e.g., 80 NADPH-consuming genes in E. coli) [4].
- Transformation: Co-transform the production strain with the dCas9* plasmid and individual sgRNA plasmids.
- Screening: Inoculate transformants into production medium in shake flasks. Monitor cell growth and product titer.
- Hit Identification: Identify strains where sgRNA repression leads to increased product yield without severely inhibiting growth. In the 4HPAA study, repression of
yahK(NADPH-consuming) andfecE(ATP-consuming) were identified as hits [4]. - Validation & Optimization: Delete or further repress the identified gene targets. For fine-tuning, implement dynamic regulation systems (e.g., quorum-sensing repressor system) to downregulate targets only at the optimal fermentation stage [4].
Performance Metrics for Cofactor Engineering
Quantitative Data from Protein Engineering Studies This table summarizes key performance indicators from a review of 103 enzyme engineering studies, providing benchmarks for your projects [38].
| Engineering Strategy | Success Rate (Specificity Reversed) | Avg. Coenzyme Specificity | Avg. Relative Catalytic Efficiency |
|---|---|---|---|
| Loop Exchange | High | >1 | Highest among strategies |
| Rational Design (Single/Double Mutations) | Moderate | >1 | Often < 0.5 |
| Directed Evolution | Variable | Variable | Can be high after optimization |
| Computational Design | Emerging | >1 | Promising, often higher than rational design |
- Key Metrics Definitions [38]:
- Coenzyme Specificity Ratio:
(kcat/Km)NADP / (kcat/Km)NADwhen switching to NADP, or vice versa. A value >1 indicates success. - Relative Catalytic Efficiency:
(kcat/Km)mutant, desired cofactor / (kcat/Km)WT, natural cofactor. This measures how much efficiency was retained.
- Coenzyme Specificity Ratio:
Visualizations
Cofactor Engineering Workflow
Cofactor Specificity Switch Logic
The Scientist's Toolkit
Research Reagent Solutions
| Item | Function/Brief Explanation | Example/Note |
|---|---|---|
| Genetically Encoded Biosensors | Enable real-time, non-destructive monitoring of intracellular ATP/ADP and NADPH/NADP+ ratios in live cells [36]. | e.g., iATPSnFR for ATP; Allows tracking of cofactor dynamics during fermentation. |
| CRISPRi/dCas9 System | Enables targeted repression (knock-down) of specific genes without complete knockout, ideal for studying essential ATP/NADPH-consuming genes [4]. | Used in CECRiS strategy to screen 80 NADPH- and 400 ATP-consuming genes in E. coli. |
| Site-Directed Mutagenesis Kits | Standard kit for introducing specific point mutations into plasmid DNA for rational protein engineering. | Commercial kits available from suppliers like NEB or Agilent. |
| Flux Balance Analysis (FBA) | A mathematical modeling approach used to simulate and analyze metabolic fluxes, predicting the effect of genetic manipulations on cofactor balances and growth [39]. | Used to elucidate condition-dependent roles of enzymes like FBPase in cofactor metabolism. |
Troubleshooting Guides
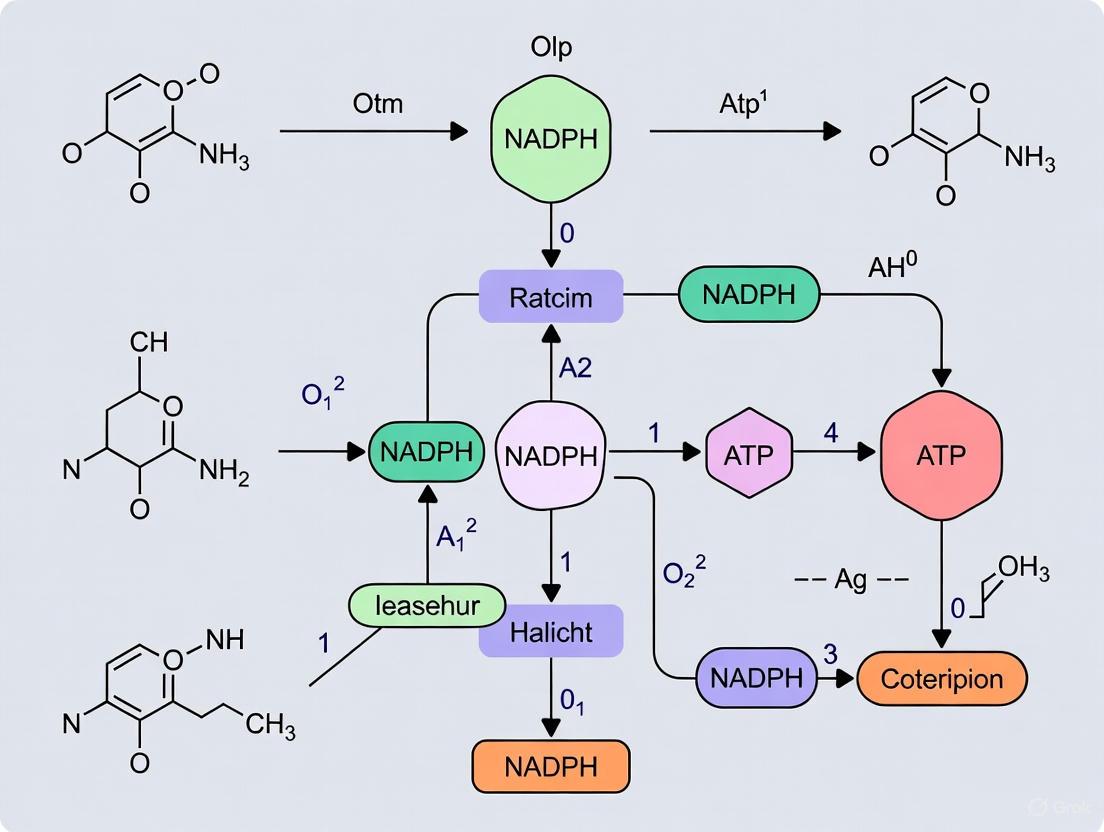
FAQ 1: How does a molecular purge valve function to maintain NADPH balance in a cell-free system?
A molecular purge valve is a synthetic biochemistry module designed to automatically maintain redox balance (NADPH/NADP⁺) in engineered pathways where cofactor generation and utilization are unbalanced. It functions by creating a metabolic node that dissipates excess reducing equivalents while maintaining carbon flux.
Core Mechanism: The system typically employs a combination of enzymes with different cofactor specificities. For example, a purge valve may use both NAD⁺-utilizing (PDHNADH) and NADP⁺-utilizing (PDHNADPH) pyruvate dehydrogenase enzymes, alongside a water-forming NADH oxidase (NoxE).
- Under Low NADPH Conditions: The NADP⁺-utilizing enzyme is more active, generating NADPH to restore balance.
- Under High NADPH Conditions: The NADP⁺-utilizing enzyme activity is choked off. The NAD⁺-utilizing enzyme takes over, producing NADH instead. The accompanying NADH oxidase (NoxE) then oxidizes this excess NADH to NAD⁺, effectively "purging" the excess reducing power and preventing a bottleneck. This allows carbon flux to continue unimpeded toward product synthesis [40] [41].
The following diagram illustrates the workflow and logical relationships within a molecular purge valve system:
FAQ 2: What are the common issues when implementing a purge valve and how can they be resolved?
| Problem | Possible Cause | Solution |
|---|---|---|
| Low Product Yield | Inefficient purge valve operation causing cofactor imbalance and metabolic bottleneck. | Optimize the ratio of PDHNADH, PDHNADPH, and NoxE enzymes. Titrate enzyme concentrations to find the optimal balance for your specific pathway [40]. |
| Enzyme Instability | Use of enzymes lacking sufficient stability for long-duration reactions. | Utilize thermostable enzymes (e.g., from Geobacillus stearothermophilus) or engineer enzymes for improved stability [40]. |
| Incomplete Cofactor Recycling | Spontaneous oxidation of NAD(P)H or suboptimal activity of the purge valve module. | Ensure the NADH oxidase (NoxE) is highly active and specific to prevent NADH buildup. The system should generate a slight excess of cofactors to account for gradual losses [40]. |
| Poor Pathway Flux | Thermodynamically unfavorable conditions or insufficient transport of substrates into synthetic compartments. | Select electron donors with favorable reduction potentials (e.g., formate, H₂). For vesicle systems, incorporate specific membrane transporters for impermeable substrates [42]. |
FAQ 3: What quantitative performance data can be expected from a functional purge valve system?
The table below summarizes key performance metrics from foundational research on molecular purge valves.
| Application | Key Enzymes in Purge Valve | Cofactor Managed | Key Outcome / Yield |
|---|---|---|---|
| Polyhydroxybutyrate (PHB) Bioplastic | PDHNADH, PDHNADPH, NoxE [40] | NADPH/NADP⁺ | Enabled robust operation of the PHB synthesis pathway from pyruvate despite inherent cofactor imbalance [40]. |
| Isoprene Production | PDHNADH, PDHNADPH, NoxE [40] | NADPH/NADP⁺ | Allowed for high-yield production (>95%) of isoprene from pyruvate via the mevalonate pathway by maintaining redox balance [40]. |
| General Synthetic Biochemistry | Engineered PDH variants, specific oxidases [40] [43] | NADPH/NADP⁺ | System provides a >90% yield for target chemicals like biodegradable plastics by effectively decoupling cofactor production from carbon flux [44]. |
Experimental Protocols
Protocol 1: Implementing a Purge Valve for PHB Production from Pyruvate
Objective: To construct a cell-free system that converts pyruvate to polyhydroxybutyrate (PHB) using a molecular purge valve to maintain NADPH balance [40].
Materials:
- Research Reagent Solutions: See the "Research Reagent Solutions" table below for essential materials.
- Buffer: Appropriate assay buffer (e.g., Tris-HCl or phosphate buffer, pH 7.0-8.0).
- Substrate: Pyruvate solution.
- Cofactors: NAD⁺, NADP⁺.
- Enzymes: Purified Pyruvate Dehydrogenase (NAD⁺-utilizing, PDHNADH), Engineered Pyruvate Dehydrogenase (NADP⁺-utilizing, PDHNADPH), NADH oxidase (NoxE), and PHB synthesis enzymes (PhaA, PhaB, PhaC).
Procedure:
- Reaction Setup: In a reaction vessel, combine the assay buffer, pyruvate (e.g., 10-50 mM), NAD⁺, and NADP⁺ at physiological concentrations.
- Enzyme Assembly: Introduce the purge valve module enzymes. A typical starting point is a 1:1 ratio of PDHNADH to PDHNADPH, with NoxE added in excess. The total enzyme concentration should be optimized for desired flux [40].
- Pathway Initiation: Add the PHB synthesis enzymes (PhaA, PhaB, PhaC) to initiate the production pathway.
- Incubation: Allow the reaction to proceed at the optimal temperature (e.g., 30-37°C) with constant mixing.
- Monitoring: Withdraw aliquots at regular intervals. Monitor PHB production using established methods (e.g., HPLC, GC-MS). Monitor NADPH/NADP⁺ ratio using spectrophotometry (absorbance at 340 nm) or genetically encoded biosensors [27] [45].
Troubleshooting Tips:
- If PHB yield is low, titrate the ratio of PDHNADH to PDHNADPH.
- If the NADPH level remains high and the reaction stalls, increase the concentration of NoxE to enhance the purge function.
Protocol 2: Engineering Cofactor Specificity of a Key Enzyme
Objective: To engineer a pyruvate dehydrogenase (PDH) to utilize NADP⁺ instead of its native NAD⁺ cofactor, a key component for creating a functional purge valve [40].
Materials:
- Structural data for the target enzyme (e.g., E3 subunit of PDH).
- Site-directed mutagenesis kit.
- Protein expression system (e.g., E. coli).
- Protein purification reagents (e.g., affinity chromatography columns).
- Enzyme activity assay reagents (pyruvate, NADP⁺, etc.).
Procedure:
- Structural Analysis: Superimpose the structure of the target enzyme's E3 subunit with a related reductase known to accept NADP⁺ (e.g., glutathione reductase). Identify residues that coordinate the additional 2'-phosphate group on NADP⁺ [40].
- Mutagenesis Design: Design point mutations to introduce key residues (e.g., substituting a serine for a basic residue like arginine) to create a favorable binding pocket for NADP⁺.
- Library Creation: Use site-directed mutagenesis to create a variant library of the PDH enzyme.
- Screening & Expression: Screen for mutants with improved NADP⁺ utilization. Express and purify the promising mutant enzyme.
- Characterization: Determine the kinetic parameters (Km, kcat) of the engineered PDH for both NAD⁺ and NADP⁺ to confirm the switch in cofactor specificity [40].
The Scientist's Toolkit: Research Reagent Solutions
| Reagent / Enzyme | Function in Purge Valve System | Key Features / Considerations |
|---|---|---|
| PDH (NAD⁺-utilizing) | Generates acetyl-CoA and NADH from pyruvate. Provides carbon flux when NADPH is sufficient. | Wild-type enzyme. Should be stable under operating conditions. |
| PDH (NADP⁺-utilizing) | Generates acetyl-CoA and NADPH from pyruvate. Restores NADPH levels when they are low. | Requires protein engineering of the E3 subunit for NADP⁺ specificity [40]. Thermostable variants (e.g., from G. stearothermophilus) are preferred. |
| NADH Oxidase (NoxE) | Oxidizes NADH to NAD⁺, dissipating excess reducing equivalents. The "purge" mechanism. | Must be specific for NADH over NADPH. A water-forming variant is ideal to avoid reactive oxygen species [40]. |
| Genetically Encoded Biosensors (e.g., NERNST, SoxR) | Enable real-time, ratiometric monitoring of the intracellular NADPH/NADP⁺ redox status. | Critical for diagnosing imbalance and validating purge valve function in vivo or in complex mixtures [27] [45]. |
| Synthetic Lipids & Vesicles | Form compartments (protocells) for encapsulating synthetic pathways, isolating reactions. | Can be engineered with pores or transporters for substrate access [42] [46]. |
Advanced Redox Regulation Concepts
Integration with Dynamic Regulation Strategies
While purge valves are powerful for static control, the next frontier involves dynamic regulation of NADPH. This can be achieved by integrating the purge valve with genetically encoded biosensors.
Mechanism: A biosensor (e.g., based on the transcription factor SoxR) can be designed to detect the intracellular NADPH/NADP⁺ ratio [27]. When the ratio becomes imbalanced, the biosensor activates the expression of a key component of the purge valve module (e.g., the NADH oxidase, NoxE), thereby closing the feedback loop and enabling real-time, autonomous control of the redox state [27].
The following diagram illustrates this advanced integrated system:
Pathway Design Principles for Innate Cofactor Balance
Troubleshooting Guides
Common Experimental Issues and Solutions
Problem: Insufficient NADPH supply limits product yield.
- Observation: Accumulation of pathway intermediates before NADPH-dependent reaction steps; reduced cell growth.
- Diagnosis: Confirm by measuring intracellular NADPH/NADP+ ratio using kits or biosensors like the NERNST biosensor [27].
- Solutions:
- Enhance PPP Flux: Overexpress genes zwf (glucose-6-phosphate dehydrogenase) and gnd (6-phosphogluconate dehydrogenase) to increase NADPH generation via the pentose phosphate pathway [27] [47].
- Introduce Alternative Enzymes: Substitute NADH-dependent enzymes in the heterologous pathway with NADPH-dependent counterparts (e.g., replacing hydroxybutyric dehydrogenase (Hbd) with acetoacetyl-CoA reductase (PhaB)) [37].
- Dynamic Regulation: Implement a biosensor-based system (e.g., SoxR) to dynamically regulate NADPH regeneration genes in response to real-time redox status [27].
Problem: NADPH/NADP+ imbalance disrupts cellular metabolism.
- Observation: Reduced biomass yield, potentially increased reactive oxygen species (ROS).
- Diagnosis: Measure overall ATP levels and key metabolites in central carbon metabolism to assess broader metabolic impact.
- Solutions:
- Fine-tune Expression: Use promoter and RBS engineering to precisely control the expression levels of NADPH-generating enzymes (e.g., zwf, gnd), avoiding excessive flux diversion [27].
- Cofactor Preference Switch: Use protein engineering to alter an enzyme's cofactor preference from NADPH to the more stable and abundant NADH, as demonstrated with the Gre2p dehydrogenase [37].
- Integrated Cofactor Engineering: Introduce a transhydrogenase system to convert excess NADPH to NADH, or fine-tune ATP synthase subunits to balance energy and redox cofactors [48].
Problem: Low activity of a heterologous, cofactor-dependent enzyme.
- Observation: The enzyme shows high expression in SDS-PAGE, but low in vivo activity, even with adequate cofactor levels.
- Diagnosis: The enzyme may be expressed as an inactive apoenzyme, lacking its essential bound cofactor (e.g., Fe-S clusters, PQQ) [49].
- Solutions:
- Co-express Cofactor Maturation Pathways: Clone and express the cofactor biosynthesis genes alongside the target enzyme. For example, express the hydE, hydF, hydG gene cluster for active Fe-Fe hydrogenase [49].
- Verify Cofactor Presence: Check for the presence of the cofactor in the host organism. For novel cofactors like PQQ, introduce the entire pqqABCDE gene cluster [49].
Problem: Inefficient Carbon Flux toward Target Product.
- Observation: High cell growth but low product titers; carbon may be diverted to biomass or by-products.
- Diagnosis: Use flux balance analysis (FBA) and (^{13}\text{C}) metabolic flux analysis to quantify flux distribution [48].
- Solutions:
- Block Competing Pathways: Knock out genes in competing pathways (e.g., pgi in glycolysis to force flux through the PPP) [47].
- Modular Pathway Engineering: Decouple pathway into growth and production modules. Use dynamic regulation to activate production modules after sufficient biomass is achieved [48].
- Synergistic Cofactor Optimization: Simultaneously engineer NADPH, ATP, and one-carbon metabolism (e.g., 5,10-MTHF) to create a balanced biosynthetic network, as demonstrated for D-pantothenic acid production [48].
Frequently Asked Questions (FAQs)
Q1: What are the primary endogenous pathways for NADPH regeneration in microbial cell factories? The main pathways are:
- Oxidative Pentose Phosphate Pathway (oxPPP): Catalyzed by glucose-6-phosphate dehydrogenase (Zwf) and 6-phosphogluconate dehydrogenase (Gnd). This is the major source [27].
- TCA Cycle: Catalyzed by NADP-dependent isocitrate dehydrogenase (Icd).
- Entner–Doudoroff (ED) Pathway: Also utilizes Zwf [27].
- NADP-dependent Malic Enzyme (Mae): Converts malate to pyruvate, generating NADPH [47].
Q2: How can I computationally design a pathway with innate cofactor balance? Tools like novoStoic use Mixed-Integer Linear Programming (MILP) to design mass-balanced pathways that simultaneously utilize known reactions and novel enzymatic steps while optimizing for cofactor balance, thermodynamic feasibility, and product yield [50]. These frameworks can embed cofactor constraints directly into the design process.
Q3: What is the difference between static and dynamic regulation strategies for cofactor balance?
- Static Regulation: Involves one-time genetic modifications, such as constitutive overexpression or knockout of genes. While simple, it can cause metabolic imbalances as it cannot adapt to changing cellular conditions [27].
- Dynamic Regulation: Uses biosensors to monitor intracellular metabolite levels (e.g., NADPH/NADP+ ratio) and dynamically regulate gene expression in response. This allows the cell to self-adjust and maintain homeostasis, leading to more robust production [27].
Q4: Why is my enzyme not active after successful heterologous expression? High expression does not guarantee functional holoenzyme formation. Many enzymes require tightly bound organic or inorganic cofactors (e.g., flavins, Fe-S clusters) for activity. You must also express the auxiliary pathways responsible for synthesizing and inserting these cofactors into the apoenzyme [49].
Q5: Can I change an enzyme's cofactor preference? Yes, through protein engineering. For instance, by mutating specific amino acids in the cofactor-binding site (e.g., Asn9 to Glu in Gre2p dehydrogenase), you can shift an enzyme's preference from NADPH to NADH, which is often more stable and cheaper to supply [37].
Quantitative Data in Pathway Engineering
Table 1: Impact of NADPH-Generating Enzyme Overexpression on Glucoamylase Production in Aspergillus niger [47]
| Overexpressed Gene | Enzyme | Pathway | Change in NADPH Pool | Effect on Glucoamylase Yield |
|---|---|---|---|---|
| gndA | 6-phosphogluconate dehydrogenase | Pentose Phosphate | +45% | +65% |
| maeA | NADP-dependent malic enzyme | Reverse TCA Cycle | +66% | +30% |
| gsdA | Glucose-6-phosphate dehydrogenase | Pentose Phosphate | Not Specified | Decrease |
Table 2: Key Cofactors and Their Roles in Microbial Biosynthesis [49] [48]
| Cofactor | Primary Role | Example in Biosynthesis |
|---|---|---|
| NADPH | Reductive biosynthesis; provides reducing power | Amino acid, lipid, and antibiotic production |
| ATP | Energy transfer; phosphorylation reactions | Active transport, polymerization reactions |
| 5,10-MTHF | One-carbon (C1) unit transfer | Precursor synthesis for nucleotides and amino acids |
| Acetyl-CoA | Acyl group carrier; central metabolic precursor | Fatty acid biosynthesis, mevalonate pathway |
Experimental Protocols
Protocol 1: Enhancing NADPH Supply via the Pentose Phosphate Pathway
Objective: Increase intracellular NADPH availability by overexpressing key PPP genes. Materials: Microbial host (e.g., E. coli, A. niger), expression plasmid, genes zwf and gnd. Methodology:
- Cloning: Amplify the zwf and gnd genes from genomic DNA. Clone them into an appropriate expression vector under a strong, inducible promoter (e.g., Ptet) [47].
- Transformation: Introduce the constructed plasmid into your production host strain.
- Cultivation: Grow the engineered strain in a defined medium. Induce gene expression at mid-exponential phase.
- Validation:
Protocol 2: Dynamic Regulation of NADPH using a Biosensor
Objective: Implement a real-time feedback system to maintain NADPH/NADP+ balance. Materials: SoxR-based biosensor for NADPH [27], inducible gene expression system. Methodology:
- Circuit Construction: Integrate the biosensor (e.g., SoxR) that controls the expression of a reporter gene (e.g., GFP) and a NADPH-regenerating gene (e.g., pos5 or zwf).
- Calibration: Characterize the biosensor response curve to different NADPH/NADP+ ratios.
- Fermentation: Cultivate the engineered strain in a bioreactor.
- Monitoring & Control: Use the biosensor's output (e.g., fluorescence) to dynamically induce the expression of NADPH-regenerating genes when the NADPH level drops below a set threshold [27].
Visualization of Workflows and Pathways
Cofactor Engineering Workflow
NADPH Regeneration Pathways
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Reagents and Tools for Cofactor Engineering
| Item | Function/Benefit | Example Use Case |
|---|---|---|
| CRISPR/Cas9 System | Enables precise genomic integration or knockout of genes. | Knocking out a competing pgi gene to redirect flux to the PPP [47]. |
| Tunable Promoters (e.g., Tet-on) | Allows precise, inducible control of gene expression levels. | Fine-tuning the expression of zwf and gnd to avoid excessive metabolic burden [47]. |
| Genetically Encoded Biosensors (e.g., SoxR, NERNST) | Enables real-time monitoring of intracellular NADPH/NADP+ ratios. | Dynamic regulation of cofactor regeneration pathways [27]. |
| NADP/NADPH Assay Kit | Quantifies the absolute levels and ratio of NADP+ and NADPH in cell extracts. | Diagnosing cofactor imbalance in engineered strains [47]. |
| Plasmids for Cofactor Maturation | Vectors containing genes for cofactor biosynthesis (e.g., hydEFG, pqqABCDE). | Activating heterologous enzymes that require complex cofactors like Fe-S clusters [49]. |
| Flux Analysis Software | In silico modeling of metabolic flux (e.g., using FBA). | Predicting flux redistribution after genetic modifications and identifying cofactor bottlenecks [48]. |
Frequently Asked Questions (FAQs)
FAQ 1: What are the most common symptoms of cofactor imbalance in an engineered pathway, and how can I diagnose them? A cofactor imbalance often manifests as incomplete substrate conversion, low product yield, or the accumulation of metabolic intermediates or by-products [33] [51]. For example, if your pathway consumes more NADPH than it is produced, you might observe the accumulation of aldehydes or acids if downstream, NADPH-dependent reactions are stalled [52]. To diagnose this:
- Measure Metabolites: Use analytics like HPLC or GC-MS to track the concentration of substrates, target products, and intermediates over time. The accumulation of an intermediate directly before an NADPH- or ATP-dependent step is a strong indicator [33].
- Analyze By-products: The appearance of side products, such as acetate in E. coli fermentations, can be a sign of redox imbalance as the cell attempts to dissipate excess reducing power [52].
- Use Computational Models: Employ Constraint-Based Analysis (CBA) or similar stoichiometric modeling techniques. These models can predict flux distributions and identify where ATP/NADPH production and consumption become uncoupled, leading to futile cycles that waste energy [33].
FAQ 2: My pathway requires NADPH, but my host mainly produces NADH. What are my options to correct this? This is a common challenge. You have two primary strategic options, which can also be combined:
- Engineer Cofactor Preference of Pathway Enzymes: Swap out the native NADPH-dependent enzymes in your heterologous pathway for isoforms that are NADH-dependent or have dual specificity. This is known as protein engineering [51] [37]. For instance, in an engineered Synechococcus elongatus strain for 1-butanol production, researchers replaced NADPH-dependent enzymes (Hbd, AdhE2) with NADH-dependent alternatives (PhaB, YqhD) to better match the host's NADPH-rich environment [37].
- Modify the Host's Cofactor Regeneration System: Re-engine the host's metabolism to increase NADPH supply. This can be done by overexpressing native transhydrogenases or introducing synthetic cycles like the Entner-Doudoroff pathway to shift the balance toward NADPH production [51] [52].
FAQ 3: How can I adjust the ATP demand in my engineered system to improve product yield? Balancing ATP is critical as both excess and deficiency can halt production [53] [33].
- For Increased ATP Demand: If your product pathway has a low ATP demand, the excess ATP may be diverted into cell growth or wasteful futile cycles, pulling resources away from your product. To counteract this, you can introduce or enhance an ATP-consuming reaction within your product pathway [33].
- For Decreased ATP Demand: If your pathway demands more ATP than the host can provide, product synthesis will stall. In such cases, you can:
- Supplement ATP-Supplying Pathways: Enhance pathways like cyclic electron flow (in photosynthetic organisms) or substrate-level phosphorylation to generate additional ATP without producing excess reducing power [53].
- Reduce Energetic Burden: De-couple high-ATP demand processes (like high-rate cell division) from the production phase by using dynamic regulatory circuits [33].
FAQ 4: What are the best practices for scaling up a cofactor-balanced pathway from lab-scale bioreactors? Scaling a cofactor-engineered strain introduces new challenges. Key considerations include:
- Maintaining Cofactor Balance Under Dynamic Conditions: Lab-scale cultures are often well-mixed and homogeneous, whereas large-scale fermenters have gradients in nutrient, oxygen, and pH. These fluctuations can disrupt the finely tuned cofactor balance achieved in the lab. It is crucial to monitor and control these parameters tightly [54].
- Economic Viability: Cofactor engineering must result in a process that is economically competitive. Utilizing cheaper cofactors like NADH over NADPH where possible, and employing robust, stable strains that maintain their engineered traits over many generations is essential for industrial application [54] [37].
- Integrated Biorefineries: Consider a biorefinery model where multiple products are generated. This can improve the overall economic and environmental footprint by utilizing different metabolic streams and balancing cofactors across parallel processes [55].
Troubleshooting Guides
Problem: Low Yield of Target Biofuel (e.g., Alkanes) in EngineeredE. coli
Background: You have introduced a synthetic alkane biosynthesis pathway (e.g., the AAR/ADO pathway) into E. coli, but the final alkane titer remains low despite good cell growth [56].
Potential Causes and Solutions:
Cause: Competition for Fatty Acid Precursors The host's native metabolism diverts fatty acyl-ACPs/CoAs away from your alkane pathway toward membrane lipid synthesis.
Cause: Inefficient Cofactor Regeneration for AAR Enzyme The cyanobacterial AAR enzyme is highly dependent on NADPH. An inadequate NADPH supply can limit the conversion of fatty acyl-ACP to fatty aldehyde.
- Solution:
- Overexpress genes in the pentose phosphate pathway (PPP), the primary source of NADPH in E. coli [51].
- Introduce a synthetic NADPH regeneration system, such as a soluble transhydrogenase [51] [52].
- Consider protein engineering to alter the cofactor specificity of AAR from NADPH to the more abundant NADH [37].
- Solution:
Cause: Toxicity or Degradation of Pathway Intermediates The fatty aldehyde intermediate can be toxic or be diverted by native host enzymes (e.g., aldehyde dehydrogenases).
- Solution: Identify and knock out native aldehyde-consuming genes. For example, in S. cerevisiae, deletion of the hexadecenal dehydrogenase gene (HFD1) increased alkane production by 22 μg/g of dry weight [56].
Experimental Protocol: Boosting Alkane Production via Cofactor Balancing
- Objective: Increase alkane titer in engineered E. coli by enhancing NADPH supply.
- Materials:
- Strain: E. coli strain engineered with AAR and ADO genes.
- Plasmids: Plasmid for overexpression of zwf (glucose-6-phosphate dehydrogenase, a key PPP enzyme).
- Growth Medium: Defined minimal medium (e.g., M9) with glucose as carbon source.
- Analytical Equipment: GC-MS for alkane quantification, HPLC for metabolite analysis.
- Methodology:
- Strain Construction: Transform the zwf overexpression plasmid into your alkane-producing E. coli strain. Include a control strain with an empty plasmid.
- Cultivation: Inoculate triplicate cultures of both strains in shake flasks. Monitor growth (OD₆₀₀) and take samples at regular intervals (e.g., every 3-4 hours).
- Sampling:
- For Alkane Titer: Extract alkanes from culture broth using an organic solvent (e.g., dodecane overlay or hexane extraction) and analyze via GC-MS [56].
- For Metabolomics: Quench cell metabolism rapidly and measure intracellular concentrations of NADPH, NADP⁺, and key central carbon metabolites to confirm the increased NADPH/NADP⁺ ratio.
- Data Analysis: Compare the maximum alkane titer and yield between the engineered and control strains. A statistically significant increase confirms that NADPH supply was a limiting factor.
Problem: Accumulation of By-products in Bioplastic (PHA) Production
Background: During the production of bioplastics like polyhydroxyalkanoates (PHA) in Cupriavidus necator, you observe significant accumulation of by-products like pyruvate or acetate, indicating metabolic imbalance [52].
Potential Causes and Solutions:
- Cause: Redox Imbalance (Excess NADH)
Under conditions of high carbon flux, an over-reduced state (high NADH) can inhibit key enzymes in the TCA cycle, causing intermediates to spill over into by-products like pyruvate and acetate.
- Solution:
- Introduce Electron Sinks: Express a synthetic, NADH-consuming pathway that does not compete with product synthesis, such as the E. coli soluble transhydrogenase (pntAB) which can convert NADH to NADPH [51].
- Modulate Aeration: Fine-tune the aeration rate in the bioreactor to regulate the oxidative phosphorylation rate, which is a major consumer of NADH.
- Solution:
Quantitative Data on Cofactor Engineering in Bio-Production
Table 1: Summary of Cofactor Engineering Strategies and Their Impact on Product Yield
| Target Product | Host Organism | Cofactor Challenge | Engineering Strategy | Outcome | Key Reference |
|---|---|---|---|---|---|
| 1-Butanol | Synechococcus elongatus (Cyanobacteria) | Host is NADPH-rich, but pathway enzymes were NADH-dependent. | Replaced NADH-dependent enzymes (Hbd, AdhE2) with NADPH-dependent ones (PhaB, YqhD). | Enabled efficient production in a cyanobacterial host by matching cofactor preference. | [37] |
| n-Alkanes (Biofuel) | Escherichia coli | NADPH limitation for the AAR enzyme in the synthetic pathway. | Overexpression of PPP genes (e.g., zwf) to boost NADPH supply. | Increased alkane titer; addressing a key cofactor limitation. | [51] [56] |
| n-Alkanes (Biofuel) | Saccharomyces cerevisiae | Native aldehyde dehydrogenases consumed the fatty aldehyde intermediate. | Deletion of hexadecenal dehydrogenase gene (HFD1). | Increased alkane titer by 22 μg/g DCW by preventing intermediate loss. | [56] |
| Isobutanol | Escherichia coli | Cofactor imbalance in the synthetic pathway limited theoretical yield. | Changed cofactor specificity of a key enzyme from NADPH to NADH. | Achieved 100% of theoretical yield by optimizing the ATP/NAD(P)H balance. | [51] |
The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential Reagents for Cofactor Balancing Experiments
| Reagent / Material | Function / Application | Example Use Case |
|---|---|---|
| Plasmids for Gene Overexpression | Vectors for introducing or modulating expression of genes involved in cofactor regeneration (e.g., pntAB, zwf). | Enhancing NADPH supply in E. coli by overexpressing the pentose phosphate pathway [51]. |
| CRISPR-Cas9 System | Tool for precise gene knock-outs or edits to remove competing pathways or regulatory elements. | Knocking out aldehyde dehydrogenase (HFD1) in yeast to prevent loss of alkane precursors [56]. |
| Heterologous Enzymes (e.g., PhaB, YqhD) | Pre-characterized enzymes with alternative cofactor specificities to replace native enzymes in a pathway. | Switching a pathway's cofactor preference from NADH to NADPH to match host metabolism [37]. |
| Computational Models (e.g., CBA, FBA) | Stoichiometric models (like the E. coli core model) to predict flux distributions and identify cofactor imbalance in silico. | Using Constraint-Based Analysis to predict and quantify ATP/NADPH imbalances in butanol production pathways [33]. |
Pathways and Workflows
Cofactor Balance Management Workflow
The diagram below outlines a systematic workflow for diagnosing and resolving cofactor imbalance in an engineered metabolic pathway.
Resolving Imbalance: Diagnostic Tools and Optimization Strategies for Enhanced Performance
Troubleshooting Guides
Guide 1: Diagnosing Cofactor Imbalance in Engineered Microbial Strains
Problem: Engineered microbial strain shows reduced growth and suboptimal target product yield despite successful pathway integration.
Explanation: This frequently indicates a cofactor imbalance where the introduced synthetic pathway disrupts the cellular redox (NAD(P)H) or energy (ATP) equilibrium. The host's native metabolism cannot adequately supply or recycle the required cofactors, causing metabolic burden and diverting flux away from product formation [16].
Diagnostic Steps:
- Measure Intracellular Cofactor Pools: Quantify NADPH/NADP+ and ATP/ADP/AMP ratios in your strain versus the control. A significantly lower ratio indicates excessive consumption by the synthetic pathway.
- Analyze Byproduct Secretion: Profile fermentation broth for accumulated metabolites like acetate, lactate, or xylitol. These byproducts are often "overflow metabolites" that the cell uses to regenerate oxidized cofactors (e.g., NAD+) when faced with redox imbalance [57] [16].
- Compute Theoretical Cofactor Demand: Calculate the net ATP and NAD(P)H consumption or generation of your heterologous pathway. Compare this to the host's central metabolic network capacity to identify stoichiometric imbalances [58] [16].
Solutions:
- Apply Cofactor Engineering: Introduce a transhydrogenase to convert NADH to NADPH, or use protein engineering to switch the cofactor preference of a key enzyme from NADPH to NADH (or vice versa) to balance the redox load [58] [57].
- Implement a Cofactor Regeneration System: Enhance the pentose phosphate pathway (PPP) flux to boost NADPH supply by overexpressing glucose-6-phosphate dehydrogenase (Zwf) and 6-phosphogluconate dehydrogenase (Gnd) [58].
- Employ Dynamic Regulation: Use sensors to dynamically regulate pathway expression in response to intracellular cofactor levels, preventing excessive burden [58].
Guide 2: Addressing Cofactor Limitations in In Vitro Biosynthetic Systems
Problem: Cell-free biosynthesis system experiences a rapid decline in reaction rate, failing to sustain long-term production.
Explanation: In vitro systems lack the regenerative machinery of a living cell. The reaction halts because key cofactors (NADPH, ATP, FAD) are consumed and not regenerated, making cofactor depletion a primary bottleneck [59].
Diagnostic Steps:
- Monitor Cofactor Depletion: Use enzymatic assays or HPLC to track the concentration of NADPH, ATP, and other cofactors over time. A steady decline confirms depletion.
- Profile Reaction Intermediates: Accumulation of pathway intermediates, particularly those upstream of a cofactor-dependent reaction step, can pinpoint where the limitation occurs.
Solutions:
- Design a Cofactor Recycling System: Incorporate complementary enzyme pairs to regenerate cofactors. For example, use glucose dehydrogenase (GDH) to regenerate NADPH from NADP+ while consuming glucose [60].
- Assemble Enzyme Complexes: Use synthetic scaffolds (e.g., peptide/protein pairs like SpyTag/SpyCatcher) to co-localize pathway enzymes with cofactor regeneration enzymes. This creates a "metabolic channel" that efficiently recycles cofactors locally, enhancing overall pathway efficiency [59].
Frequently Asked Questions (FAQs)
FAQ 1: What are the most common metabolic signatures of NADPH imbalance?
The table below summarizes key diagnostic markers for NADPH imbalance.
Table: Metabolic Signatures and Diagnostic Markers for Cofactor Imbalance
| Marker Category | Specific Signature | Underlying Cause / Interpretation |
|---|---|---|
| Metabolic Byproducts | Accumulation of xylitol in pentose sugar fermentation [57] | Imbalance between NADPH-preferred XR and NAD+-preferred XDH in fungal D-xylose pathway. |
| Secretion of acetate or lactate [16] | Regeneration of NAD+ from NADH due to redox imbalance. | |
| Flux Analysis | Reduced flux through the oxidative Pentose Phosphate Pathway (PPP) [58] | Inability to meet high NADPH demand, leading to pathway bottleneck. |
| Theoretical Calculation | Negative net NADPH balance in the synthetic pathway [16] | Pathway consumes more NADPH than the host's central metabolism can produce. |
FAQ 2: What computational tools can predict cofactor imbalance before experimental implementation?
Genome-scale metabolic models (GEMs) and constraint-based analysis are key tools.
- Flux Balance Analysis (FBA): Used to predict metabolic flux distributions and identify if a proposed pathway creates an unsustainable demand for ATP or NAD(P)H, which the model may "solve" by producing unrealistic futile cycles or excess biomass instead of the target product [16] [26].
- Cofactor Balance Analysis (CBA): A specialized algorithm used with FBA to track and categorize how ATP and NAD(P)H pools are affected by a new pathway, helping to quantify the imbalance [16].
- Dynamic FBA (DFBA): Simulates batch fermentations over time, predicting the temporal utilization of substrates and the formation of products, which can reveal cofactor-related bottlenecks during different fermentation phases [57].
FAQ 3: How can I experimentally monitor real-time cofactor levels in live cells?
Autofluorescence of metabolic cofactors can be exploited for non-destructive, real-time monitoring.
- NAD(P)H and FAD Fluorescence: NAD(P)H and FAD are intrinsically fluorescent. Their fluorescence intensity and lifetime can be measured using two-photon excited fluorescence (TPEF) microscopy or flow cytometry [61] [62].
- Optical Redox Ratio: The ratio of FAD to NAD(P)H fluorescence intensity (FAD/(NAD(P)H+FAD)) provides a sensitive measure of the cellular redox state. A lower ratio can indicate a more reduced state, potentially signaling imbalance [61].
- Fluorescence Lifetime Imaging (FLIM): FLIM measures the average time a fluorophore spends in the excited state. The lifetime of NAD(P)H is sensitive to whether it is free or enzyme-bound, providing insights into metabolic activity beyond what intensity alone can show [61].
FAQ 4: Can cofactor imbalance ever be beneficial for production?
Yes, a novel strategy known as Redox Imbalance Forces Drive (RIFD) deliberately creates an excess NADPH state to drive metabolic flux toward a target product.
- Principle: By combining "open source" (enhancing NADPH supply) and "reduce expenditure" (knocking out non-essential NADPH-consuming genes) strategies, an imbalanced driving force is created. The cell then utilizes the product-forming pathway as a "sink" to alleviate this redox pressure, thereby restoring balance and increasing product yield [58]. This approach has successfully enhanced L-threonine production in E. coli [58].
The Scientist's Toolkit: Essential Research Reagents and Materials
Table: Key Reagents for Investigating Cofactor Balance
| Reagent / Material | Function / Application |
|---|---|
| Two-Photon Microscope | Enables high-resolution, non-invasive imaging of NAD(P)H and FAD autofluorescence in live cells and tissues for metabolic assessment [61]. |
| Glucose-6-Phosphate Dehydrogenase (Zwf) | A key enzyme in the oxidative PPP; its overexpression is a common "open source" strategy to increase intracellular NADPH pools [58]. |
| Transhydrogenases (e.g., PntAB, UdhA) | Enzymes that facilitate conversion between NADH and NADPH pools, used to modulate cofactor specificity and balance redox state [58]. |
| Xylose Reductase (XR) & Xylitol Dehydrogenase (XDH) | Enzymes in the fungal D-xylose pathway; a classic model system for studying NADPH/NAD+ imbalance and a target for cofactor specificity engineering [57]. |
| SpyTag/SpyCatcher Peptide-Protein Pair | A synthetic biology tool for creating self-assembled protein scaffolds. Used to co-localize metabolic pathway enzymes to substrate channel and improve cofactor recycling efficiency [59]. |
| Genome-Scale Metabolic Model (e.g., iMM904, E. coli core model) | Computational framework for in silico prediction of metabolic fluxes and cofactor demands of engineered pathways using FBA and CBA [57] [16]. |
Visualizing Workflows and Pathways
RIFD Strategy Workflow
Cofactor Balance Analysis with FBA
ATP Waste Mechanisms and Substrate Cycles for Energy Dissipation
Frequently Asked Questions (FAQs)
1. What is a substrate cycle (futile cycle) and why would a cell 'waste' ATP? A substrate cycle occurs when two metabolic pathways run simultaneously in opposite directions, resulting in no net conversion of substrate but dissipating energy as heat. While historically termed "futile," these cycles are now understood to be critical metabolic regulators. They provide a mechanism for sensitive control of metabolite concentrations, generate heat for thermal homeostasis (e.g., in brown adipose tissue of young mammals or in insect flight muscles), and can enhance lipolysis and energy homeostasis at the whole-body level [63]. In the context of metabolic engineering, they can be engineered to help maintain cofactor balance, such as the NADPH/NADP+ balance [64].
2. How can substrate cycles be exploited in metabolic engineering? In engineered pathways, activating specific substrate cycles can be a strategy to manage cofactor levels. For instance, research has shown that activating the pyruvate-phosphoenolpyruvate (PEP) futile cycle in skeletal muscle can enhance lipolysis in adipose tissues and orchestrate crosstalk between tissues to control whole-body energy homeostasis [63]. Furthermore, in an engineered E. coli strain, a metabolic route towards acetol biosynthesis was triggered under nitrogen limitation, which proved favorable for maintaining the NADPH/NADP+ balance during production, making product formation mandatory for the cell's cofactor equilibrium [64].
3. What are common issues when managing ATP levels in engineered microbes? A primary challenge is unbalanced overall metabolic homeostasis. Reconstituting pathways for high-efficiency chemical production often disturbs the intracellular availability and dynamic balance of essential cofactors like ATP, NADPH, and others [48]. This can lead to redox imbalance, energy deficits, and the accumulation of toxic intermediates, ultimately restricting the metabolic flux toward the desired product. Strategies to address this include fine-tuning ATP synthase subunits rather than simply overexpressing them, and introducing heterologous systems to convert excess reducing equivalents (NADPH, NADH) into ATP [48].
4. What experimental methods are used to quantify ATP and NADPH in cultures? A common methodology involves HPLC-UV analysis of extracted cofactors [64]. The general workflow is:
- Rapid Sampling & Quenching: A sample of cell broth is quickly mixed with perchloric acid in an overhead shaker at 4°C. The acidic conditions stabilize oxidized cofactors (NAD+, NADP+) while preventing inaccuracies from the oxidation of reduced forms [64].
- Neutralization: The sample is neutralized using K₂HPO₄ and KOH while kept in ice water [64].
- Analysis: The neutralized supernatant is analyzed via HPLC-UV to quantify the energy and redox cofactors [64].
5. How is intracellular metabolic flux analyzed? ¹³C-flux analysis is a powerful technique for elucidating flux re-routing in central carbon metabolism. In practice:
- Cells are fed a carbon source with a defined ¹³C-label, such as 2-¹³C glycerol [64].
- The incorporation of the labeled carbon into intracellular metabolites and proteinogenic amino acids is tracked [64].
- This labeling data is used to calculate the intracellular flux distribution under different conditions (e.g., exponential growth vs. nitrogen starvation), revealing how the cell redirects its metabolic resources in response to genetic or environmental perturbations [64].
Troubleshooting Guide
| Problem | Potential Cause | Suggested Solution |
|---|---|---|
| Low product yield despite high pathway gene expression | Inefficient cofactor regeneration leading to redox/energy imbalance [48]. | Implement an integrated cofactor engineering strategy. Enhance NADPH regeneration by screening endogenous/heterologous genes (e.g., gapN) [48]. Fine-tune ATP synthase activity and introduce a transhydrogenase to convert excess NADPH to ATP [48]. |
| Unbalanced NADPH/NADP+ ratio | High demand for NADPH in biosynthetic pathways without sufficient recycling mechanisms [64]. | Engineer a futile cycle or a compensating pathway. For example, activate the pyruvate-PEP futile cycle or introduce an acetol biosynthesis pathway that consumes NADPH to restore balance under non-growth conditions [63] [64]. |
| Accumulation of metabolic intermediates | Insufficient ATP supply or feedback inhibition disrupting the pathway [65] [48]. | Modulate allosteric regulation. Overexpress enzymes that are insensitive to ATP feedback inhibition (e.g., certain PFK-1 mutants). Use FBA/FVA to identify and relieve ATP-mediated negative feedback loops on key enzymes like pyruvate kinase [65] [48]. |
| Poor cell growth and low ATP | Metabolic burden from heterologous pathway expression draining cellular ATP [48]. | Decouple growth from production. Use nutrient limitation (e.g., nitrogen) to trigger a non-growing production state. Optimize carbon flux through EMP/PPP/ED pathways via flux balance analysis to ensure adequate ATP and precursor supply [48] [64]. |
Quantitative Data on Substrate Cycles
The table below summarizes key substrate cycles and their energy dissipation characteristics, based on current research.
| Substrate Cycle | Organism / System | Net Reaction | Primary Physiological Role | Key Enzymes Involved |
|---|---|---|---|---|
| Glycolysis / Gluconeogenesis (F6P/F1,6BP node) | General metabolism | ATP + H₂O → ADP + Pi + Heat [63] | Metabolic regulation, heat generation [63] | Phosphofructokinase-1 (PFK-1), Fructose-1,6-bisphosphatase (FBPase-1) [63] |
| Pyruvate-PEP Futile Cycle | Skeletal muscle (via miR-378) | Consumption of ATP, activation enhances lipolysis [63] | Regulates whole-body energy homeostasis, ameliorates obesity in mouse models [63] | Pyruvate carboxylase, PEP carboxykinase (PEPCK), and others |
| Glycolysis / Gluconeogenesis | Zebrafish swim bladder | ATP + H₂O → ADP + Pi + Heat [63] | Critical heat generation for gas gland cell function and buoyancy control [63] | Fructose-1,6-bisphosphatase (Fbp1), Glyceraldehyde-3-phosphate dehydrogenase (Gapdh) [63] |
| Glycolysis / Gluconeogenesis | Bumblebee flight muscle | ATP + H₂O → ADP + Pi + Heat [63] | Rapid heat generation for warming up bodies at low ambient temperatures [63] | Fructose-1,6-bisphosphatase (Fbp), Phosphofructokinase (Pfk) [63] |
Detailed Experimental Protocols
Protocol 1: Analyzing Cofactor Levels via HPLC-UV
Objective: To quantify intracellular concentrations of ATP, ADP, NADPH, and NADP+ from microbial cultures.
Materials:
- Quenching Solution: 20-25% (v/v) Perchloric acid [64]
- Neutralization Solutions: 1 M K₂HPO₄, 5 M KOH [64]
- HPLC System: Equipped with a UV detector [64]
- Chromatography Column: LiChrospher RP-18 (25 cm length, 4.6 mm diameter) [64]
- Centrifuge: Capable of maintaining 4°C and achieving 4,696 x g [64]
Procedure:
- Sampling & Quenching: Rapidly collect 4 mL of cell broth and immediately mix it with 1 mL of ice-cold perchloric acid. Incubate in an overhead shaker for 15 minutes at 4°C to fully quench metabolism and lyse cells [64].
- Neutralization: Transfer the quenched sample to an ice-water bath. Gradually add appropriate volumes of 1 M K₂HPO₄ and 5 M KOH to neutralize the sample to a pH of ~6.5-7.5. Monitor the pH to avoid overshooting [64].
- Clarification: Centrifuge the neutralized sample at 4,696 x g for 10 minutes at 4°C to pellet cell debris. Carefully collect the supernatant [64].
- Storage: Aliquot and store the clarified supernatant at -20°C until HPLC analysis [64].
- HPLC-UV Analysis: Inject the sample into the HPLC system. Use a validated gradient method with two buffers to separate the cofactors. Identify and quantify them by comparing retention times and peak areas to those of known standards [64].
Protocol 2: ¹³C-Metabolic Flux Analysis (¹³C-MFA)
Objective: To determine intracellular metabolic flux distributions in engineered strains under different conditions.
Materials:
- Labeled Substrate: e.g., 2-¹³C Glycerol (99% atom purity) [64]
- Bioreactor: Controlled for pH, temperature, and dissolved oxygen (e.g., BioFlo 3000) [64]
- GC-MS or LC-MS: For measuring isotopic labeling in metabolites [64]
Procedure:
- Strain Cultivation: Grow the engineered strain (e.g., E. coli) in a defined minimal medium with the natural (unlabeled) carbon source. Use a bioreactor for controlled environmental conditions [64].
- Isotope Labeling Pulse: Once the desired culture phase is reached (e.g., nitrogen limitation for production phase), introduce the ¹³C-labeled carbon source (e.g., 2-¹³C glycerol) into the medium [64].
- Metabolite Sampling: Rapidly sample the culture at multiple time points after the pulse. Quench metabolism immediately (e.g., using cold methanol). Extract intracellular metabolites [64].
- Mass Spectrometry Analysis: Derivatize and analyze the extracted metabolites (e.g., amino acids from protein hydrolysis) via GC-MS or LC-MS to determine the mass isotopomer distributions [64].
- Flux Calculation: Use specialized software (e.g., INCA, OpenFlux) to fit the experimental labeling data to a metabolic network model. The software will iteratively adjust metabolic fluxes until the simulated labeling patterns match the measured data, providing a quantitative map of intracellular flux [64].
Pathway and Workflow Visualizations
Diagram Title: Pyruvate-PEP Futile Cycle Logic
Diagram Title: Acetol Production Under Nitrogen Limitation
The Scientist's Toolkit: Research Reagent Solutions
| Reagent / Material | Function in Research | Application Example |
|---|---|---|
| 2-¹³C Glycerol | A stable isotope-labeled carbon source for tracing metabolic fate of carbon atoms and quantifying intracellular fluxes via ¹³C-MFA [64]. | Used to elucidate flux re-routing towards acetol biosynthesis in E. coli during nitrogen starvation [64]. |
| MitoTracker Dyes | Cell-permeant fluorescent probes that accumulate in active mitochondria, enabling visualization and quantification of mitochondrial localization and morphology [66]. | Used to demonstrate mitochondrial re-localization to the nuclear periphery under mechanical cell confinement [66]. |
| Perchloric Acid / KOH | Metabolite quenching and neutralization agents. Perchloric acid rapidly halts metabolic activity; KOH neutralizes the extract for stable analysis of acid-stable cofactors [64]. | Essential for the accurate quantification of ATP, NADPH, and other cofactors via HPLC-UV in E. coli cultures [64]. |
| HPLC-UV System with LiChrospher RP-18 Column | Analytical platform for separating and quantifying cofactors and other metabolites based on their retention times and UV absorption [64]. | Employed for the simultaneous measurement of energy and redox cofactors (ATP, NADPH) in cell extracts [64]. |
| Heterologous Transhydrogenase System (e.g., from S. cerevisiae) | An engineered enzyme system that can convert excess reducing equivalents (NADPH and NADH) into ATP, helping to couple and balance redox and energy metabolism [48]. | Integrated into E. coli to form a redox-energy coupling strategy, enhancing production of D-pantothenic acid [48]. |
Troubleshooting Common NADPH Engineering Problems
Q1: I have engineered the Pentose Phosphate Pathway (PPP) in my microbial cell factory, but my product yield remains low and cell growth is impaired. What could be wrong?
A: This is a common issue when the carbon flux is excessively diverted toward NADPH production at the cost of essential growth metabolites. The problem likely involves carbon economy imbalance.
- Root Cause: The oxidative PPP generates NADPH but also releases CO₂, leading to carbon loss [67] [47]. Over-reliance on this pathway can deplete the cell of carbon skeletons (like ribose-5-phosphate) needed for nucleotide synthesis and aromatic amino acids, thereby inhibiting growth and overall productivity.
- Solution: Consider implementing a dual-strategy:
- Partially restore glycolytic flux: Avoid complete knockout of PPP entry points. Instead, use tunable promoters to fine-tune the expression of genes like ZWF1 (glucose-6-phosphate dehydrogenase) [67] [3].
- Introduce carbon-efficient alternatives: Express heterologous NADP⁺-dependent glyceraldehyde-3-phosphate dehydrogenase (GAPDH). This enzyme redirects carbon flux from the PPP back into glycolysis while simultaneously generating NADPH, without the carbon loss associated with the oxidative PPP [67] [47] [68]. This strategy has been shown to increase ethanol yield in Saccharomyces cerevisiae by 13.5% and improve production of compounds like L-lysine [67] [47].
Q2: My product requires high NADPH, but standard PPP engineering is not providing sufficient cofactor supply. What are effective alternative NADPH regeneration strategies?
A: When the native PPP is insufficient, you can engineer "bypass" pathways to enhance NADPH supply. The most effective strategies are summarized in the table below.
Table 1: Alternative NADPH Regeneration Pathways for Metabolic Engineering
| Strategy | Key Enzyme(s) | Mechanism | Example Application & Result |
|---|---|---|---|
| Cofactor Specificity Switching | Engineered NADP⁺-dependent GAPDH [67] [68] | Converts glycolytic NADH production to NADPH production. | In E. coli and Y. lipolytica, this redirects central carbon flux, increasing yields of lysine and β-carotene without carbon loss [47] [68]. |
| Transhydrogenase Cycles | Native or heterologous transhydrogenases (e.g., UdhA, PntAB) [69] [58] | Reversibly transfers hydride ions between NADH and NADP⁺ pools. | Useful for balancing overall redox state. In E. coli, manipulation of UdhA and PntAB can influence NADPH-dependent product synthesis [58]. |
| NAD(H) Kinase Expression | POS5 (NADH kinase) [70] [3] | Phosphorylates NADH to generate NADPH directly. | Expression of cPOS5 from S. cerevisiae in P. pastoris increased α-farnesene production by improving the NADPH supply [3]. |
| Malic Enzyme Pathway | NADP-dependent malic enzyme (MAE) [47] | Decarboxylates malate to pyruvate, generating NADPH. | Overexpression of maeA in Aspergillus niger increased the intracellular NADPH pool by 66% and glucoamylase yield by 30% [47]. |
Q3: How can I systematically identify and resolve NADPH limitations in a non-model organism without a well-annotated genome?
A: A multi-faceted "DBTL" (Design-Build-Test-Learn) approach is recommended to diagnose and address cofactor limitations.
- Diagnostic "Test" Phase:
- Metabolomics: Measure intracellular concentrations of NADP⁺ and NADPH, and the ratios of key PPP metabolites (e.g., sedoheptulose-7-phosphate) to infer pathway activity [71] [47].
- ¹³C Metabolic Flux Analysis (¹³C-MFA): This is the gold standard for quantifying the in vivo flux through the PPP versus glycolysis. An insufficient flux through the oxidative PPP will be directly visible [47].
- Intervention "Build" Phase:
- Heterologous Expression: Introduce and test a panel of conserved, well-characterized NADPH-generating enzymes (e.g., GAPDH from Clostridium acetobutylicum, NADH kinases) on plasmids with inducible promoters [47] [68].
- Complementation Assays: Knock down a native NADPH-generation gene (e.g., ZWF1). The ability of candidate heterologous genes to restore growth on a defined medium indicates their functional efficacy in your host [71] [67].
Quantitative Comparison of NADPH Generation Systems
Choosing the right NADPH regeneration strategy requires a balance of cofactor yield, carbon efficiency, and impact on host metabolism. The following table provides a quantitative comparison based on experimental data.
Table 2: Performance Metrics of Engineered NADPH Regeneration Systems
| Engineering Strategy | Host Organism | Target Product | Impact on NADPH Pool / Product Titer | Reported Key Metric |
|---|---|---|---|---|
| Overexpression of PPP Enzymes (gndA) | Aspergillus niger | Glucoamylase (GlaA) | ↑ NADPH pool by 45%; ↑ GlaA yield by 65% [47] | 65% increase in protein yield [47] |
| NADP⁺-GAPDH Replacement | Saccharomyces cerevisiae | Ethanol (from xylose) | ↑ Ethanol yield by 13.5% [67] | 1.6-fold increase in xylose consumption rate [67] |
| NADH Kinase (POS5) Expression | Pichia pastoris | α-Farnesene | ↑ α-Farnesene production by 41.7% [3] | Final titer of 3.09 g/L in shake flasks [3] |
| Malic Enzyme (maeA) Overexpression | Aspergillus niger | Glucoamylase (GlaA) | ↑ NADPH pool by 66%; ↑ GlaA yield by 30% [47] | 30% increase in protein yield [47] |
| Redox Imbalance Drive (RIFD) | Escherichia coli | L-Threonine | ↑ L-Threonine titer to 117.65 g/L [58] | Yield of 0.65 g/g glucose [58] |
Detailed Experimental Protocols
Protocol: Inverting Cofactor Specificity of a Key Glycolytic Enzyme
This protocol is adapted from a structure-guided, semi-rational strategy for reversing the cofactor specificity of oxidoreductases [18].
1. Structural Analysis and Library Design:
- Input Structure: Obtain a high-resolution crystal structure of your target enzyme (e.g., GAPDH) in complex with its native cofactor (NAD⁺).
- Identify Specificity-Determining Residues: Use the web tool CSR-SALAD (Cofactor Specificity Reversal - Structural Analysis and LibrAry Design) to identify residues within ~6 Å of the 2'-phosphate moiety of NADP⁺ [18]. These typically include residues that bind the adenosine ribose and those that can be mutated to create a favorable electrostatic environment for the 2'-phosphate.
- Design a Degenerate Codon Library: CSR-SALAD will propose a set of "sub-saturation" degenerate codons to mutagenize the targeted residues. This approach keeps the library size experimentally tractable (e.g., 10³-10⁵ variants) by focusing on structurally similar amino acid substitutions known to facilitate cofactor switches [18].
2. Library Construction and Primary Screening:
- Clone the Library: Generate the mutant library using site-saturation mutagenesis or gene synthesis and clone it into an appropriate expression vector.
- High-Throughput Activity Assay: Express the library in a 96-well plate format. Lyse cells and assay for enzymatic activity using the non-preferred cofactor (NADP⁺). A colorimetric or fluorescent assay linked to NADPH production is ideal for rapid screening.
- Isolate Hits: Select the top 0.5-1% of variants showing the highest activity with NADP⁺ for further characterization.
3. Characterization and Activity Recovery:
- Secondary Screening: Purify the hit variants and determine their kinetic parameters (Kₘ, k꜀ₐₜ) for both NAD⁺ and NADP⁺ to quantify the specificity reversal.
- Combinatorial Mutagenesis: Combine beneficial mutations from different hits to further improve catalytic efficiency with NADP⁺.
- Address Activity Loss: Cofactor-switched enzymes often suffer from reduced activity. Use structure-guided mutagenesis to target residues around the adenine ring of the cofactor to recover stability and activity [18].
Protocol: Implementing a Heterologous NADPH Regeneration Pathway in Yeast
This protocol outlines the steps for integrating and testing the phosphoketolase (PK)-phosphotransacetylase (PTA) pathway or a NADP⁺-GAPDH in Saccharomyces cerevisiae or Yarrowia lipolytica [68].
1. Strain and Plasmid Construction:
- Gene Selection: Codon-optimize and synthesize genes for your chosen pathway. For the PK-PTA pathway, use xfp (phosphoketolase) and pta (phosphotransacetylase) from Aspergillus nidulans or other suitable sources. For GAPDH replacement, use gapN (NADP⁺-dependent GAPDH) from Clostridium acetobutylicum [68].
- Vector Assembly: Clone these genes into a yeast expression vector under the control of strong, constitutive promoters (e.g., PTEF1, PGPD). Include selectable markers (e.g., LEU2).
- Host Transformation: Introduce the expression vector into your production strain using standard lithium acetate transformation. Select positive transformants on appropriate dropout media.
2. Metabolic Flux Validation:
- Culture Conditions: Grow the engineered and control strains in defined minimal medium with labeled [1-¹³C] glucose.
- Metabolite Extraction and Analysis: Harvest cells at mid-exponential phase. Quench metabolism rapidly and extract intracellular metabolites.
- GC-MS Analysis: Derivatize the metabolites and analyze by GC-MS. The labeling pattern in sedoheptulose-7-phosphate (a PPP intermediate) versus lower glycolysis intermediates will reveal the redirection of carbon flux [68].
3. Phenotypic Characterization:
- Growth Curve Analysis: Monitor OD₆₀₀ over time in shake flasks to assess any impact on growth kinetics.
- Product Titer Quantification: At the end of fermentation, extract and quantify your target product (e.g., β-carotene via spectrophotometry, lipids via GC, etc.). Compare the titer and yield to the control strain to determine the success of the engineering strategy [68].
Essential Visualizations
NADPH Regeneration Pathways in Microbial Cell Factories
This diagram illustrates the primary native and engineered pathways for NADPH regeneration within the central carbon metabolism of a typical microbial cell factory, highlighting key engineering targets.
Decision Workflow for NADPH Pathway Engineering
This flowchart provides a logical sequence for diagnosing NADPH limitations and selecting the most appropriate engineering strategy.
The Scientist's Toolkit: Key Research Reagents
Table 3: Essential Reagents for NADPH Cofactor Engineering
| Reagent / Tool | Function / Description | Example Use Case |
|---|---|---|
| CRISPR-Cas9 System | Enables precise gene knock-outs, knock-ins, and edits. | Essential for creating chassis strains (e.g., knocking out ZWF1 or native GAPDH) and integrating expression cassettes [47] [68]. |
| Tet-On Gene Switch | A tunable promoter system activated by doxycycline (DOX). | Allows for precise, temporal control over the expression of NADPH-generating genes (e.g., gndA, maeA) to avoid metabolic burden during early growth phases [47]. |
| CSR-SALAD Web Tool | A structure-guided, semi-rational design tool for reversing enzyme cofactor specificity. | Used to design mutant libraries for switching NAD⁺-dependent enzymes (e.g., GAPDH, Aldehyde dehydrogenase) to NADP⁺-dependent ones [18]. |
| NADP⁺-GAPDH (gapN/gapC) | Heterologous enzymes that generate NADPH during glycolysis. | Key for constructing carbon-efficient NADPH regeneration systems in E. coli, C. glutamicum, and yeasts [67] [68]. |
| NADH Kinase (POS5) | Phosphorylates NADH to generate NADPH. | Expressed heterologously in P. pastoris and S. cerevisiae to augment the NADPH pool from the NADH generated in catabolism [70] [3]. |
| Capillary Electrophoresis Mass Spectrometry (CE-MS) | Analytical platform for measuring intracellular metabolite concentrations. | Used to quantify NADP⁺/NADPH ratios and intermediates like sedoheptulose-7-phosphate to validate PPP flux [71]. |
Futile Cycle Prevention and Network-Wide Cofactor Balancing
Troubleshooting Guides and FAQs
Frequently Asked Questions
Q1: What are the primary consequences of cofactor imbalance in an engineered metabolic pathway? A primary consequence is the accumulation of intermediates and reduced product yield. For instance, in S. cerevisiae engineered with fungal D-xylose utilization pathways, a cofactor imbalance between NADPH-preferring xylose reductase (XR) and NAD+-preferring xylitol dehydrogenase (XDH) leads to significant xylitol accumulation, reducing ethanol productivity [57]. This imbalance forces the cell's metabolic network to expend energy to rebalance the NADPH/NADP+ and NADH/NAD+ pools, diverting resources away from product synthesis.
Q2: What are futile cycles and how can they be beneficial in metabolic engineering? A futile cycle occurs when two metabolic pathways run simultaneously in opposite directions, consuming ATP without performing net metabolic work, thereby dissipating energy as heat [63]. While often considered wasteful, they can be harnessed beneficially. Enforced ATP futile cycling can increase metabolic flux, drive substrate uptake, and shift carbon allocation from biomass to product formation. In E. coli, implementing an ATP futile cycle between phosphoenolpyruvate (PEP) and pyruvate increased anaerobic lactate yield by 8% and specific productivity by 25% [72].
Q3: What biochemical mechanisms exist in nature to prevent NADPH overproduction and imbalance? Bacteria use several mechanisms to balance catabolic NADPH formation with anabolic demand [6]:
- Transhydrogenases: Enzymes like the membrane-bound PntAB or soluble UdhA can directly counterbalance NADPH over- or under-production by catalyzing the reversible transfer of reducing equivalents between NADH and NADPH [6] [57].
- Dual Cofactor Specificity of Enzymes: Many catabolic enzymes, such as glucose-6-phosphate dehydrogenase, can use both NAD+ and NADP+, allowing the pathway flux to automatically adjust to the cellular redox state [6].
- Expression of Isoenzymes: Possessing multiple enzymes that catalyze the same reaction but with different cofactor specificities provides a regulatory mechanism to maintain cofactor balance [6].
Q4: My pathway requires NADPH, but my host cell seems to have insufficient supply. What are my engineering options? You can engineer the host to enhance NADPH availability [73] [57]:
- Rewrite Pathway Cofactor Specificity: Use protein engineering to change the cofactor preference of key enzymes in your pathway from NADH to NADPH, or vice-versa, to match supply and demand [73] [57].
- Express NADH Kinases: Introduce enzymes that convert NADH to NADPH at the expense of ATP, directly linking the two pools [6].
- Modulate Native Pathways: Enhance flux through the oxidative pentose phosphate pathway, a major NADPH source, or express external transhydrogenases [73].
Common Experimental Issues and Solutions
| Problem | Underlying Cause | Troubleshooting Solution | Key Performance Indicator |
|---|---|---|---|
| Low Product Yield & Intermediate Accumulation | Cofactor imbalance in heterologous pathway (e.g., XR-NADPH/XDH-NAD+) leading to redox conflict [57]. | Re-engineer enzyme cofactor specificity (e.g., XDH to NADP+); introduce soluble transhydrogenase; optimize carbon flux [57]. | Reduced intermediate (e.g., xylitol) concentration; ≥20% increase in target product yield [57]. |
| Poor Cell Growth & Metabolite Production | Energetic stress from uncontrolled, native futile cycling (e.g., simultaneous glycolysis/gluconeogenesis) [63]. | Implement dynamic regulatory control to separate opposing pathways; fine-tune enzyme expression levels rather than knockout [74]. | Improved specific growth rate; restoration of ATP pool. |
| Insufficient Metabolic Driving Force | Lack of ATP consumption, leading to low substrate uptake and glycolytic flux under anaerobic conditions [72]. | Introduce an enforced ATP futile cycle (e.g., PEP synthase/Pyk cycle) to consume ATP and pull flux [72]. | 10-25% increase in specific substrate uptake and product productivity rates [72]. |
| Low NADPH Availability | High demand from engineered pathway exceeds host's innate supply capacity [73] [57]. | Overexpress NADH kinase; modulate Pentose Phosphate Pathway (PPP) flux; engineer NADP+-dependent isoenzymes in central metabolism [73] [57]. | Increased NADPH/NADP+ ratio; elimination of NADPH-limited growth arrest. |
Essential Experimental Protocols
Protocol 1: Implementing an ATP Futile Cycle to Enhance Lactate Production inE. coli
This protocol is adapted from a study demonstrating increased yield and specific productivity of anaerobic lactate production [72].
1. Objective: To construct an IPTG-inducible ATP futile cycle between phosphoenolpyruvate (PEP) and pyruvate to increase lactate yield and productivity in a high-producing E. coli strain.
2. Materials:
- Strain: E. coli KBM10111 (or equivalent, e.g., MG1655 ΔadhE ΔackA-pta) as the high-lactate producer base strain [72].
- Plasmid: Construct an expression plasmid (e.g., pET or pBAD derivative) containing the ppsA gene (encoding PEP synthase) under the control of an IPTG-inducible promoter (e.g., PT7-lac or Ptac).
- Growth Media: M9 minimal medium supplemented with appropriate antibiotic and carbon source (e.g., 10 g/L glucose).
3. Methodology:
- Strain Transformation: Transform the ppsA expression plasmid into the E. coli KBM10111 host strain.
- Anaerobic Batch Fermentation:
- Inoculate 5 mL of aerobic pre-culture and grow overnight.
- Use this to inoculate a main anaerobic culture in a sealed bioreactor or flask with an inert atmosphere (e.g., N2).
- Grow cultures at 37°C with constant agitation.
- Induce ppsA expression by adding a predetermined optimal concentration of IPTG (e.g., 0.1 - 1.0 mM) at the early exponential growth phase (OD600 ~0.4-0.6).
- Analytical Measurements:
- Monitor cell density (OD600) periodically.
- Analyze concentrations of glucose, lactate, and any by-products (e.g., acetate, succinate) using HPLC or equivalent methods.
- Calculate specific productivity, yield, and substrate uptake rate from the time-course data.
4. Expected Outcome: Upon induction of ppsA, the engineered strain should exhibit an approximate 8% higher lactate yield and a 25% higher specific lactate productivity compared to the uninduced control [72].
Protocol 2: Computational Prediction of Cofactor Balancing Outcomes using a Genome-Scale Model
This protocol uses constraint-based modeling to predict the global metabolic impacts of cofactor balancing before undertaking laboratory work [57].
1. Objective: To use a genome-scale metabolic model (GEM) and Dynamic Flux Balance Analysis (DFBA) to predict the growth and production benefits of balancing a cofactor-imbalanced pentose utilization pathway.
2. Materials:
- Software: A constraint-based modeling software suite (e.g., COBRA Toolbox for MATLAB or Python).
- Model: A relevant GEM (e.g., S. cerevisiae iMM904 [57]).
- Reaction List: Stoichiometric definitions of the imbalanced and balanced versions of your target pathway.
3. Methodology:
- Model Modification:
- Add the reactions for the cofactor-imbalanced fungal D-xylose and L-arabinose pathways to the base GEM. This involves adding XR (NADPH-dependent) and XDH (NAD+-dependent) reactions.
- Create a second, "balanced" model where the cofactor specificity of XDH is changed from NAD+ to NADP+.
- Simulation Setup:
- Set the simulation conditions for a batch culture with mixed sugars (e.g., 20 g/L glucose, 50 g/L D-xylose, 30 g/L L-arabinose) and a microaerobic oxygen uptake rate (e.g., 0.3 mmol/gDCW/h) [57].
- Implement catabolite repression constraints to ensure glucose is consumed first.
- DFBA Simulation:
- Run DFBA simulations for both the imbalanced and balanced models to predict the time profiles of sugar consumption, biomass growth, and ethanol production.
- Analysis:
- Compare the total simulation time, final ethanol titer, and yield between the two models.
4. Expected Outcome: The simulation for the cofactor-balanced model predicts a 24.7% increase in ethanol production and a 70% reduction in the time required to utilize the substrate mixture compared to the imbalanced model [57]. This provides a quantitative justification for the enzyme engineering effort.
Pathway and Workflow Visualizations
Diagram Title: Cofactor Engineering Workflow
Diagram Title: Engineered ATP Futile Cycle for Lactate
The Scientist's Toolkit: Research Reagent Solutions
| Research Reagent | Function in Cofactor Balancing |
|---|---|
| Soluble Transhydrogenase (UdhA) | Catalyzes the energy-independent transfer of reducing equivalents between NADH and NADPH, directly addressing cofactor imbalances [6] [57]. |
| Membrane-Bound Transhydrogenase (PntAB) | Uses the proton motive force to drive reduction of NADP+ by NADH, helping to maintain a high NADPH/NADP+ ratio [6]. |
| NADH Kinase | Phosphorylates NADH to generate NADPH, providing a direct link between the NADH and NADPH pools at the cost of ATP [6]. |
| PEP Synthase (PpsA) | A key enzyme for constructing enforced ATP futile cycles. Catalyzes the ATP-dependent conversion of pyruvate to PEP [72]. |
| Genome-Scale Metabolic Model (GEM) | A computational model used to predict the systemic consequences of pathway engineering, including cofactor demand and growth phenotypes, prior to lab work [57] [75]. |
| Protein Engineering Kits | Site-directed mutagenesis kits and related reagents are essential for altering the cofactor specificity of native enzymes (e.g., changing XDH from NAD+ to NADP+ preference) [73] [57]. |
Host Selection and Chassis Engineering for Optimal Cofactor Metabolism
A technical guide for resolving NADPH and ATP imbalances in engineered microbial systems.
This technical support center provides a curated knowledge base for researchers addressing the critical challenge of cofactor balance in engineered metabolic pathways. The following guides and protocols are designed to help you diagnose and resolve issues related to NADPH and ATP management in microbial chassis.
Troubleshooting Guides
FAQ 1: Why is my engineered pathway producing low yields despite high gene expression?
This typically indicates an intracellular cofactor imbalance where the demand for reducing power or energy exceeds the host's native supply capacity.
- Problem: Low target metabolite production despite successful pathway integration and high enzyme expression levels.
- Underlying Cause: Cofactor imbalance, particularly insufficient NADPH supply, creating a bottleneck in the metabolic flux. The heterologous pathway competes with host maintenance for essential cofactors, disrupting redox homeostasis [76] [47].
- Diagnosis:
- Monitor for auxiliary metabolite secretion (e.g., xylitol accumulation in pentose sugar pathways) indicating redox imbalance [57].
- Measure intracellular NADPH/NADP+ ratios using enzymatic assays or biosensors.
- Perform metabolic flux analysis to identify flux bottlenecks.
- Solutions:
- Enhance NADPH regeneration by overexpressing key pentose phosphate pathway enzymes (glucose-6-phosphate dehydrogenase, 6-phosphogluconate dehydrogenase) [77] [47].
- Implement transhydrogenase cycles to convert NADH to NADPH [48] [78].
- Rewrite central carbon metabolism to create synthetic NADPH regeneration cycles [78].
FAQ 2: How do I select the optimal host for my cofactor-intensive pathway?
Host selection critically influences cofactor availability due to inherent differences in native metabolic networks and regulatory systems.
- Problem: Suboptimal production stems from mismatched cofactor demand and host supply capabilities.
- Diagnosis: Evaluate host physiology against pathway requirements:
- Bacterial systems (E. coli): Prefer NADH-dependent metabolism; require engineering for NADPH-dependent pathways [76] [48].
- Yeast systems (S. cerevisiae, P. pastoris): Naturally support NADPH-dependent pathways with eukaryotic protein processing; generally recognized as safe (GRAS) [76] [57].
- Filamentous fungi (A. niger): Robust native NADPH generation but complex background metabolism [76] [47].
- Solutions:
Table 1: Host Organism Comparison for Cofactor Metabolism
| Host Organism | Advantages for Cofactor Engineering | Limitations | Ideal Application |
|---|---|---|---|
| E. coli | Extensive genetic tools; Fast growth; Well-characterized central metabolism [48] | Primarily NADH-dependent; Limited eukaryotic enzyme compatibility [76] | Bacterial pathways; High-growth production systems |
| S. cerevisiae | Eukaryotic cofactor metabolism; GRAS status; Compartmentalization [76] [57] | Lower diversity of native secondary metabolites [76] | Eukaryotic pathways; NADPH-intensive products |
| P. pastoris | Strong, regulated promoters; High-density cultivation [76] | Methanol metabolism required for some promoters [76] | Secreted proteins; Scalable production |
| A. niger | High native NADPH generation; Industrial enzyme production [47] | Complex background metabolism; Slower growth [76] | Fungal natural products; High-titer metabolites |
FAQ 3: How can I dynamically regulate cofactor levels without impairing host growth?
Static overexpression of cofactor-generating enzymes often creates metabolic burdens; dynamic regulation provides a superior solution.
- Problem: Engineered cofactor pathways disrupt central metabolism, reducing host growth and fitness.
- Underlying Cause: Rigid metabolic engineering creates irreversible flux imbalances, draining resources from essential cellular functions [48].
- Solutions:
- Implement tunable expression systems (e.g., Tet-on, inducible promoters) to fine-tune cofactor enzyme levels [47].
- Develop biosensor-regulated systems that respond to intracellular NADPH/ATP levels.
- Engineer cofactor redundancy by creating multiple parallel regeneration systems [48].
- Utilize dynamic metabolic flux control to balance EMP, PPP, and TCA pathways [48].
Experimental Protocols
Protocol 1: Enhancing NADPH Regeneration in E. coli
This protocol details steps to improve NADPH availability for NADPH-dependent pathways in bacterial systems, adapted from high-efficiency D-pantothenic acid production strategies [48].
- Objective: Increase intracellular NADPH pool size and regeneration capacity.
- Key Reagents:
- Plasmid systems for NADPH regeneration enzyme expression
- Chromosomal integration components (CRISPR/Cas9, lambda Red)
- Analytical standards for NADPH quantification
- Procedure:
- Overexpress NADPH-generating enzymes:
- Modify cofactor specificity:
- Engineer ketopantoate reductase (PanE) and ketol-acid reductoisomerase (IlvC) to accept both NADH and NADPH [48].
- Reduce NADPH consumption:
- Delete NADPH-consuming redundant pathways (e.g., sthA) to minimize wasteful NADPH oxidation [48].
- Implement transhydrogenase systems:
- Introduce soluble transhydrogenase (UdhA) to enable NADH to NADPH conversion [48].
- Validation:
- Measure NADPH/NADP+ ratio using spectrophotometric assays.
- Quantify target metabolite yield and byproduct formation.
- Analyze growth characteristics to ensure minimal fitness impact.
Table 2: Cofactor Engineering Strategies and Outcomes
| Engineering Strategy | Specific Approach | Reported Outcome | Application Example |
|---|---|---|---|
| PPP Enhancement | Overexpress G6PDH (gsdA) and 6PGDH (gndA) [47] | 9-fold NADPH increase in A. niger; 65% GlaA yield increase [47] | Glucoamylase production [47] |
| Cofactor Specificity Switching | Change XDH cofactor preference from NAD+ to NADP+ [57] | 24.7% increase in predicted ethanol production [57] | Pentose fermentation in yeast [57] |
| Transhydrogenase Cycle | Implement GDH1/GDH2 cycle or UdhA [78] | Rescued Δpgi1 growth defect; Enhanced reducing power [78] | Synthetic reductive metabolism [78] |
| ATP-NAD(P)H Coupling | Engineer NADH-to-ATP conversion systems [48] | Improved energy charge and product yield [48] | D-pantothenic acid production [48] |
| One-Carbon Metabolism | Enhance 5,10-MTHF supply via serine-glycine cycle [48] | Supported hydroxymethylation steps in D-PA biosynthesis [48] | Cofactor-intensive vitamin production [48] |
Protocol 2: Balancing NADPH and ATP Supply in Yeast
This protocol enables coordinated management of redox and energy cofactors in eukaryotic systems, based on synthetic reductive metabolism approaches [78].
- Objective: Create balanced redox and energy metabolism for reduced chemical production.
- Key Reagents:
- Yeast integration plasmids
- CRISPR/Cas9 components for yeast
- Metabolite analysis kits (ATP, NADPH, glycerol, succinate)
- Procedure:
- Construct PP Cycle for NADPH Generation:
- Combine oxidative and non-oxidative PP pathways with gluconeogenesis to create a cyclic NADPH regeneration system [78].
- Implement Transhydrogenase Cycle:
- Co-express GDH1 (NADP-dependent) and GDH2 (NAD-dependent) to convert NADPH to NADH [78].
- Enhance ATP Supply:
- Block Competing Pathways:
- Delete pgi1 to redirect flux from glycolysis to PP pathway [78].
- Modify TCA cycle to align with synthetic energy metabolism goals.
- Construct PP Cycle for NADPH Generation:
- Validation:
- Monitor free fatty acid production (theoretical yield up to 40%) [78].
- Measure growth rates under different energy source conditions.
- Quantify glycerol and succinate production as indicators of reductive metabolism.
The Scientist's Toolkit
Table 3: Essential Research Reagents for Cofactor Engineering
| Reagent / Tool | Function | Application Examples |
|---|---|---|
| Tet-On Gene Switch | Tunable gene expression system [47] | Controlled overexpression of NADPH-generating enzymes in A. niger [47] |
| Soluble Transhydrogenase (UdhA) | Converts NADH to NADPH [48] | Redox balancing in E. coli and yeast [48] |
| NADP-GAPDH (GapCcae) | Generates NADPH in glycolysis [48] | Redirecting glycolytic flux for NADPH generation [48] |
| CRISPR/Cas9 Systems | Precision genome editing [47] | Gene knockouts, promoter replacements, pathway integration [47] |
| Genome-Scale Metabolic Models | Predict metabolic flux and cofactor demand [76] [57] | In silico testing of engineering strategies [57] |
| Biosensors | Monitor intracellular cofactor levels | Dynamic regulation of cofactor pathways |
Metabolic Pathway Diagrams
Cofactor Balancing in Pentose Utilization Pathways
Host Selection and Engineering Workflow
Measuring Success: Analytical Techniques and Comparative Assessment of Engineering Strategies
Genetically Encoded Biosensors for Real-Time NADPH/NADP+ Monitoring
The NADPH/NADP+ redox couple is a central metabolic hub, providing reducing power for anabolic pathways and antioxidative defense. In the context of engineered metabolic pathways, maintaining the balance of this cofactor is paramount, as it directly influences the yield and efficiency of biosynthesis for compounds ranging from pharmaceuticals to biofuels. An imbalance can lead to pathway bottlenecks, oxidative stress, and cell toxicity. Genetically encoded biosensors represent a revolutionary tool for researchers, allowing for real-time, non-destructive monitoring of the NADPH/NADP+ ratio with subcellular resolution in live cells. This technical support center provides a comprehensive guide to employing these biosensors, addressing common challenges, and integrating their use into the broader goal of managing cofactor balance in metabolic engineering.
Understanding the Tools: A Guide to NADPH/NADP+ Biosensors
A significant advancement in this field is the development of the NAPstar family of biosensors. These sensors enable specific, real-time monitoring of the NADP redox state (NADPH/NADP+ ratio) across a wide dynamic range.
NAPstar Biosensor Family Characteristics
The table below summarizes the key properties of different NAPstar variants to help you select the most appropriate sensor for your experimental needs.
| NAPstar Variant | Kr (NADPH/NADP+) | Kd (NADPH) (μM) | Key Characteristics & Recommended Use |
|---|---|---|---|
| NAPstar1 | ~0.006 | 0.9 | Highest affinity for NADPH; ideal for detecting low ratios or in compartments with low total NADP pool. |
| NAPstar2 | Information Missing | 2.0 | Balanced affinity; a good general-purpose starting point for cytosolic measurements. |
| NAPstar3 | ~0.03 | 2.8 | Common choice for in vivo applications; offers a solid balance of sensitivity and dynamic range. |
| NAPstar6 | ~0.08 | 11.6 | Lower affinity; suitable for environments with highly reduced NADP pools or higher total NADP. |
| NAPstar7 | Information Missing | 9.3 | Lower affinity; similar applications to NAPstar6. |
| NAPstarC | Non-functional | Non-functional | Crucial negative control; contains mutations that prevent nucleotide binding. |
Source: Data adapted from NAPstar characterization studies [79] [80].
Biosensor Mechanism and Workflow
The NAPstars are based on a circularly permuted T-Sapphire (cpTS) fluorescent protein inserted between two NADP-binding bacterial Rex domains. Binding of NADPH or NADP+ induces a conformational change that alters the fluorescence of cpTS. The signal is normalized using a constitutively fluorescent mCherry (mC) protein fused to the construct, allowing for ratiometric measurement that is independent of sensor concentration.
Troubleshooting Guides and FAQs
FAQ 1: My biosensor shows no signal or very low signal. What could be wrong?
Answer: A low or absent signal can stem from several issues. Follow this diagnostic checklist:
- Check Expression and Health: Confirm that your biosensor is expressing in the target cells using the mCherry (or other reference FP) channel. Ensure that transfection/transduction was successful and that cells are viable, as promoter strength and cell health directly impact signal intensity [81].
- Verify Subcellular Localization: If you are using a targeted biosensor (e.g., mitochondrial), confirm its correct localization using high-resolution microscopy. Mislocalization can render the sensor non-functional or place it in a compartment with a pH outside the sensor's usable range [81].
- Confirm Instrumentation Setup: Ensure your microscope or plate reader is configured with the correct excitation and emission filters. For NAPstars, you need capabilities for ~400/515 nm (cpTS) and ~570/610 nm (mCherry). Check laser power and detector sensitivity [81] [79].
FAQ 2: The biosensor signal is unstable or drifting over time. How can I stabilize it?
Answer: Signal drift is often related to environmental factors or sensor performance.
- Control for pH Artifacts: Genetically encoded sensors can be sensitive to pH fluctuations. Always perform parallel control experiments expressing cpVenus or the sensor's fluorescent protein scaffold alone to account for pH-induced fluorescence changes. The NAPstars were designed for limited pH sensitivity, but this control remains critical [81] [79].
- Ensure Buffer and Media Stability: Use well-buffered culture media (e.g., containing 25 mM HEPES) during live imaging, especially outside a CO₂ incubator, to prevent drifts in extracellular pH that can influence intracellular pH [81] [82].
- Check for Photobleaching and Toxicity: Minimize light exposure and use low illumination intensities where possible. Perform control experiments to rule out that the observed drift is due to light-induced cellular stress or photobleaching of the sensor [82].
FAQ 3: The sensor response seems inconsistent or does not match expected physiological changes.
Answer: This can be due to calibration issues, sensor saturation, or off-target effects.
- Perform a Positive Control Test: Validate that your sensor is functioning correctly by applying known perturbations. Treat cells with 1-10 mM DTT (a reducing agent) to increase the NADPH/NADP+ ratio, or with 100-500 μM H₂O₂ (an oxidizing agent) to decrease it. A lack of response indicates a problem with the sensor, expression, or measurement setup [79] [83].
- Confirm Sensor Operating Range: Ensure the NADPH/NADP+ ratio in your experimental system is within the dynamic range of your chosen NAPstar variant (see table above). If the ratio is too high or too low, the sensor signal may be saturated or minimally responsive [79] [80].
- Verify Specificity: The NAPstars are highly specific for the NADP couple over the NAD couple, but it is good practice to confirm that your experimental manipulations are primarily affecting the intended redox couple. Correlate biosensor readings with endpoint biochemical assays if possible [79].
FAQ 4: How do I convert the ratiometric signal into a quantitative NADPH/NADP+ value?
Answer: Quantitative conversion requires an in-situ calibration curve.
- In-Vitro Calibration (Recommended for rigorous quantification): Purify the biosensor protein and measure its fluorescence ratio across a range of known NADPH/NADP+ ratios in a cuvette or plate reader. This generates a standard curve to which your cellular data can be fitted [79].
- In-Cello Calibration: At the end of your live-cell experiment, permeabilize the cells and perfuse them with buffers containing defined NADPH/NADP+ ratios (e.g., using a mixture of NADPH and NADP+ with a redox mediator like glutathione). This accounts for the intracellular environment and allows you to define the minimum (Rmin) and maximum (Rmax) ratio values for your specific cells [81] [83].
- Report Ratio of Ratios: For many applications, reporting the normalized biosensor ratio (e.g., F/F₀) is sufficient to illustrate dynamic changes in the NADP redox state without absolute quantification [81].
Detailed Experimental Protocol: Measuring NADPH/NADP+ Dynamics in an Engineered Pathway
This protocol outlines the key steps for using NAPstar biosensors to monitor cofactor balance in your metabolic engineering projects.
Workflow: From Sensor Expression to Data Analysis
Step 1: Construct Preparation and Validation
- Clone your chosen NAPstar variant (e.g., NAPstar3) into an appropriate mammalian expression vector with a promoter suitable for your cell line (e.g., CMV, EF1α). For compartment-specific analysis, add targeting sequences (e.g., mitochondrial targeting signal) to the construct [79] [83].
- Critical Control: Always prepare a parallel construct of the non-binding control, NAPstarC, to account for non-specific effects.
Step 2: Cell Culture and Biosensor Expression
- Culture your engineered cell line (e.g., HEK293T, CHO, or a custom industrial cell line) under standard conditions [81].
- Transfect cells with the NAPstar plasmid DNA using a method optimized for your cell line (e.g., lipofection, electroporation). For long-term studies, consider generating stable cell lines.
- Allow 24-48 hours for robust biosensor expression before imaging. Confirm expression and subcellular localization using confocal microscopy.
Step 3: Live-Cell Imaging Setup
- Prior to the experiment, replace the culture medium with a pre-warmed, CO₂-independent imaging medium (e.g., Leibovitz's L-15 medium or phenol-red free DMEM with 25 mM HEPES, pH 7.4) [82].
- Mount the cell culture dish on a confocal or widefield microscope equipped with environmental control (37°C).
- Set up acquisition for simultaneous or sequential ratiometric imaging:
- Excitation for cpTS: 405 nm and 488 nm lasers.
- Emission for cpTS: 500-540 nm bandpass filter.
- Excitation for mCherry: 560 nm laser.
- Emission for mCherry: 590-650 nm bandpass filter.
Step 4: Experimental Perturbation and Data Acquisition
- Acquire a stable baseline for at least 5-10 minutes.
- Introduce your experimental perturbation:
- Pathway Induction: Add an inducer (e.g., doxycycline for a synthetic pathway) to initiate product synthesis and drain NADPH.
- Substrate Feeding: Add a carbon source (e.g., glucose, xylose) to stimulate NADPH generation.
- Inhibitor/Drug Treatment: Add compounds that target pathways linked to NADPH metabolism (e.g., GSH synthesis inhibitors).
- Continue time-lapse imaging for the duration required to capture the dynamic response, typically 30 minutes to several hours.
Step 5: Data Analysis
- Use image analysis software (e.g., ImageJ/Fiji, CellProfiler) to segment cells and measure background-subtracted mean fluorescence intensities in both the cpTS (F₅₁₅) and mCherry (F₆₁₀) channels for each cell and time point.
- Calculate the ratiometric value (R) for each cell: R = F₅₁₅ / F₆₁₀.
- Normalize the data as needed, for example, as F/F₀, where F₀ is the average baseline ratio.
The Scientist's Toolkit: Essential Research Reagents
This table lists key materials and their functions for successfully implementing NADPH/NADP+ biosensing experiments.
| Reagent / Material | Function / Application | Examples / Notes |
|---|---|---|
| NAPstar Plasmids | Genetically encoded sensor for NADPH/NADP+ ratio. | NAPstar1-7, NAPstarC (control); available from Addgene or original authors. |
| Cell Line | Cellular host for biosensor expression and pathway engineering. | HEK293T [81], CHO, iPSC-derived cells, or custom industrial strains. |
| Transfection Reagent | Introducing biosensor DNA into cells. | Lipofectamine, polyethylenimine (PEI), or electroporation systems. |
| CO₂-Independent Medium | Maintains pH during live-cell imaging outside an incubator. | Leibovitz's L-15 or FluoroBrite DMEM supplemented with HEPES [81] [82]. |
| Chemical Perturbagens | Positive controls for sensor validation and experimental tools. | DTT (reducer), H₂O₂ (oxidizer), pathway-specific substrates/inhibitors. |
| Confocal/Microscope System | High-resolution, time-lapse ratiometric imaging. | Systems from Zeiss, Nikon, Olympus, or equivalent, with 405/488/561 nm lasers. |
Integrating Biosensor Data with Cofactor Balance Management
The ultimate goal of using these biosensors is to inform and guide metabolic engineering strategies. A decreased NADPH/NADP+ ratio in your engineered strain upon pathway induction clearly indicates a cofactor bottleneck. This data can direct you to implement solutions such as:
- Overexpressing NADPH-Generating Enzymes: Introduce genes for glucose-6-phosphate dehydrogenase (G6PD), malic enzyme, or NADP+-dependent isocitrate dehydrogenase.
- Engineering Transhydrogenases: Implement synthetic cycles to shuttle reducing equivalents from the NADH to the NADPH pool.
- Optimizing Pathway Flux: Use biosensor data as a real-time feedback signal to dynamically control pathway expression, avoiding excessive metabolic burden and maintaining redox homeostasis.
By providing a window into the real-time dynamics of central redox metabolism, genetically encoded biosensors like the NAPstars are indispensable tools for diagnosing and resolving cofactor imbalance, thereby accelerating the development of robust and efficient engineered biological systems [79] [83].
Flux Balance Analysis and Metabolic Flux Determination for Cofactor Utilization Assessment
Flux Balance Analysis (FBA) is a mathematical approach for analyzing the flow of metabolites through a metabolic network, particularly genome-scale metabolic reconstructions [84]. This constraint-based method calculates the steady-state flux distributions in metabolic networks, enabling researchers to predict biological outcomes such as cellular growth rates or the production of biotechnologically important metabolites without requiring extensive kinetic parameter data [84] [85].
For researchers engineering pathways with cofactor balance requirements, FBA provides a computational framework to simulate how manipulations affect NADPH and ATP utilization. The method operates on the principle that metabolic systems reach a steady state where metabolite concentrations remain constant, allowing the formulation of mass-balance constraints using stoichiometric matrices [85]. This makes FBA particularly valuable for predicting how engineered changes impact the delicate balance of energy cofactors in cellular systems.
Key Principles and Mathematical Foundation
FBA is built upon constraint-based modeling principles that differentiate it from traditional kinetic modeling approaches [84]. The core mathematical framework represents the metabolic network as a stoichiometric matrix (S) where rows correspond to metabolites and columns represent metabolic reactions [84].
The fundamental equation governing FBA is: Sv = 0 where S is the stoichiometric matrix and v is the flux vector of all reaction rates in the network [84] [85]. This equation represents the steady-state assumption that metabolite concentrations remain constant over time.
FBA utilizes linear programming to identify flux distributions that maximize or minimize a specified objective function (Z = cᵀv), which represents the biological goal of the system [84] [85]. For cofactor utilization studies, this typically involves maximizing biomass production while analyzing the resulting fluxes through NADPH- and ATP-producing or consuming reactions.
Table 1: Key Components of FBA Mathematical Framework
| Component | Symbol | Description | Application in Cofactor Studies |
|---|---|---|---|
| Stoichiometric Matrix | S | m × n matrix mapping metabolites to reactions | Defines cofactor participation in reactions |
| Flux Vector | v | n × 1 vector of reaction rates | Quantifies cofactor production/consumption rates |
| Objective Function | Z = cᵀv | Linear combination of fluxes to optimize | Often biomass formation; can be NADPH/ATP yield |
| Constraints | lb ≤ v ≤ ub | Lower and upper flux bounds | Limits cofactor exchange rates |
Troubleshooting FBA for Cofactor Studies
Why does my FBA predict unrealistic cofactor production?
This common issue often stems from incomplete constraint application. FBA may exploit network gaps to generate energy and cofactors without appropriate biological costs [86]. The Metabolite Dilution FBA (MD-FBA) approach addresses this by accounting for growth-associated dilution of all intermediate metabolites, including cofactors [86].
Solution: Apply additional constraints to cofactor cycling reactions based on experimental measurements. Implement MD-FBA to prevent unrealistic cofactor recycling without synthesis costs. For catalytic cofactors like ATP and NADPH, ensure your model includes:
- Maintenance energy requirements
- Realistic bounds on exchange fluxes
- Compartment-specific constraints where applicable
How can I improve gene essentiality predictions for cofactor-related genes?
Traditional FBA may mispredict gene essentiality because it doesn't account for cofactor dilution during growth [86]. In one study, MD-FBA correctly identified 12% more essential genes in E. coli compared to standard FBA by properly accounting for metabolic costs of cofactor synthesis [86].
Solution:
- Use single gene deletion analysis by constraining the flux through gene-associated reactions to zero [85]
- Implement MD-FBA to account for dilution of cofactors
- Validate predictions with experimental data on cofactor auxotrophy
Why do my simulations show inconsistent ATP/NADPH production across conditions?
This often results from missing network elements or incorrect biomass composition. FBA models commonly use a constant biomass composition across conditions, but actual cofactor demands vary with genetic background and growth media [86].
Solution:
- Verify your model includes all known cofactor utilization routes
- Check for catalytic cycles that might be artificially generating cofactors
- Perform phenotypic phase plane analysis to understand how cofactor production shifts with nutrient availability [85]
Experimental Protocols for Cofactor-FBA Integration
Protocol 1: Validating FBA Predictions with Cofactor Measurements
Purpose: To experimentally verify predicted NADPH/ATP flux distributions.
Materials:
- Wild-type and engineered microbial strains
- LC-MS system for cofactor quantification
- Bioreactor with controlled conditions
- Quenching solution (60% methanol, -40°C)
Procedure:
- Cultivate strains in biologically relevant media under controlled conditions
- Collect samples at mid-exponential phase for intracellular metabolite extraction
- Quench metabolism rapidly using cold methanol solution
- Extract and quantify NADPH, NADP+, ATP, ADP using LC-MS
- Compare absolute cofactor concentrations and ratios to FBA predictions
- Iteratively refine model constraints based on discrepancies
Protocol 2: Gene Knockout Analysis for Cofactor Pathway Validation
Purpose: To test FBA predictions of gene essentiality in cofactor metabolism.
Materials:
- Gene knockout library or CRISPR-Cas9 system
- Selective growth media
- Microplate reader for growth phenotyping
Procedure:
- Select target genes involved in cofactor metabolism based on FBA predictions
- Create knockout strains using appropriate genetic tools
- Measure growth phenotypes across multiple nutrient conditions
- Compare experimental results with FBA predictions of essentiality
- Use discrepancies to identify missing network elements or incorrect constraints
Table 2: Essential Research Reagents for Cofactor-FBA Studies
| Reagent/Resource | Function | Example Application |
|---|---|---|
| Genome-scale metabolic model | Network representation | iJO1366 (E. coli), Yeast8 (S. cerevisiae) |
| COBRA Toolbox | FBA computation | Performing flux balance analysis in MATLAB [84] |
| LC-MS instrumentation | Cofactor quantification | Measuring intracellular ATP/NADPH concentrations |
| CRISPR-Cas9 system | Genetic manipulation | Creating knockout mutants for model validation |
| Phenotypic microarray | High-throughput growth assays | Testing growth under various conditions |
Computational Workflows and Visualization
FBA Workflow for Cofactor Balance Assessment
Figure 1: FBA workflow for assessing cofactor utilization in metabolic networks.
Cofactor Cycling in Metabolic Networks
Figure 2: ATP and NADPH coupling in metabolic networks for biomass production.
Advanced FBA Techniques for Cofactor Engineering
Metabolite Dilution FBA (MD-FBA)
Standard FBA fails to account for growth-associated dilution of intermediate metabolites, including cofactors, which can lead to biologically implausible predictions [86]. MD-FBA addresses this limitation by incorporating dilution terms for all metabolites produced in the network.
Implementation:
- Formulated as a mixed-integer linear programming (MILP) problem
- Accounts for dilution of catalytic cofactors in metabolic cycles
- Particularly important for cofactors like ATP and NADPH that participate in multiple reactions
In application to E. coli models, MD-FBA demonstrated improved prediction of gene essentiality and growth rates across different conditions by properly accounting for the metabolic costs of cofactor synthesis [86].
Flux Variability Analysis (FBA) for Cofactor Pathways
FVA determines the range of possible fluxes through each reaction while maintaining optimal objective function value [84]. For cofactor studies, this helps identify:
- Alternative pathways for cofactor production
- Flexibility in cofactor utilization
- Potential engineering targets for redirecting cofactor fluxes
Frequently Asked Questions
How accurate are FBA predictions for NADPH/ATP balance?
FBA predictions have shown good correlation with experimental measurements, but accuracy depends on model quality and constraint appropriateness. Studies comparing FBA predictions with experimental growth rates under different conditions have demonstrated reasonable agreement, though predictions for cofactor-specific fluxes may require additional validation [84] [86]. MD-FBA shows improved accuracy by accounting for metabolite dilution effects [86].
What are the limitations of FBA for cofactor studies?
Key limitations include:
- Lack of regulatory information (allosteric regulation, phosphorylation)
- Inability to predict metabolite concentrations
- Steady-state assumption prevents dynamic analysis
- Dependence on accurate biomass composition
- Potential gaps in cofactor-related reactions in network reconstructions
How can I integrate regulatory constraints for cofactor metabolism?
While standard FBA doesn't include regulation, you can:
- Add flux bounds based on known regulatory patterns
- Use rFBA (regulatory FBA) if regulatory network information is available
- Implement dFBA (dynamic FBA) for time-varying simulations
- Incorporate proteomic constraints to account for enzyme expression costs
What objective functions are most appropriate for cofactor studies?
The choice depends on your research question:
- Biomass maximization for simulating natural selection
- ATP/NADPH production for studying energy metabolism
- Product yield for metabolic engineering applications
- Non-growth associated maintenance for stationary phase studies
Flux Balance Analysis provides a powerful framework for assessing cofactor utilization in engineered pathways, but requires careful implementation to produce biologically relevant predictions. By addressing common troubleshooting scenarios, applying appropriate experimental validation protocols, and utilizing advanced methods like MD-FBA, researchers can significantly improve the reliability of cofactor balance predictions in their metabolic engineering projects.
Troubleshooting Guide: Cofactor Imbalance in Engineered Pathways
FAQ: Low Product Yields and Growth Issues
Q1: Our engineered strain shows low product yield despite high substrate consumption. Metabolic flux analysis suggests a redox imbalance. What are the primary investigative steps?
A: This pattern often indicates insufficient NADPH regeneration capacity. Follow this investigative protocol:
- Quantify Cofactor Pools: Measure intracellular NADPH/NADP+ and ATP/ADP ratios using LC-MS techniques as described in recent E. coli riboflavin studies [87].
- Analyze Competing Pathways: Use Flux Balance Analysis (FBA) to identify NADPH-consuming reactions in central metabolism that may be draining reducing equivalents [48].
- Profile Pathway Intermediates: Measure concentrations of key pathway metabolites to identify thermodynamic bottlenecks where low driving forces may cause flux limitations [88].
Experimental Protocol 1: Quantifying Intracellular Cofactor Pools
- Principle: Extract and quantify adenosine and nicotinamide nucleotides via liquid chromatography-mass spectrometry.
- Procedure:
- Rapid Quenching: Culture samples are rapidly quenched in 60% methanol at -40°C.
- Metabolite Extraction: Use 80% ethanol extraction with 0.1 M formic acid for nucleotide stabilization.
- LC-MS Analysis: Separate on a HILIC column with a mobile phase of ammonium acetate and acetonitrile. Detect using negative ion mode MRM.
- Quantification: Calculate NADPH/NADP+ and ATP/ADP ratios from standard curves using isotopically labeled internal standards [48].
Q2: Pathway simulations predict thermodynamic feasibility, but in vivo flux remains low. How can we identify and resolve kinetic bottlenecks?
A: This discrepancy suggests kinetic limitations despite thermodynamic permissibility. Implement this multi-objective optimization approach:
- Calculate Minimum Driving Force (MDF): Use optimization tools to compute the MDF across your pathway, identifying steps with smallest ΔG margins [88].
- Evaluate Flux-Force Efficacy (FFE): Apply the FFE formula to quantify enzyme utilization efficiency: FFE = (e^(ΔGdiss/RT) - 1) / (e^(ΔGdiss/RT) + 1), where a higher FFE indicates better forward flux [88].
- Screen Enzyme Variants: Test orthologous enzymes from thermophilic or alternative hosts that may operate more efficiently under your pathway conditions [89].
Q3: We need to enhance NADPH availability for our D-pantothenic acid production strain. What engineering strategies have demonstrated success?
A: Recent studies show integrated approaches work best:
- Enhance NADPH Regeneration: Overexpress glucose-6-phosphate dehydrogenase (Zwf) and a soluble transhydrogenase (PntAB) to increase pentose phosphate pathway flux [48].
- Introduce Heterologous Systems: Implement a non-phosphorylating glyceraldehyde-3-phosphate dehydrogenase (GapN) to generate NADPH directly in glycolysis [48].
- Dynamic TCA Cycle Regulation: Use CRISPRi to downregulate citrate synthase (gltA) during production phase, redirecting carbon from NADH-generating TCA toward NADPH-generating pathways [48].
Experimental Protocol 2: Flux Variability Analysis for Cofactor Balancing
- Principle: Use constraint-based modeling to identify flux distributions that satisfy cofactor demands while maximizing product yield.
- Procedure:
- Model Construction: Build a genome-scale metabolic model incorporating your heterologous pathway.
- Constraint Definition: Set substrate uptake rate and maintenance ATP requirements as constraints.
- FVA Implementation: Perform Flux Variability Analysis to determine the feasible range of each reaction flux while maximizing NADPH availability.
- Gene Knockout Prediction: Identify gene deletion targets that minimize competitive NADPH consumption while maintaining product formation [48].
Quantitative Metrics for Pathway Evaluation
Table 1: Thermodynamic and Kinetic Efficiency Metrics for Pathway Analysis
| Metric | Calculation Formula | Optimal Range | Application in Pathway Design |
|---|---|---|---|
| Minimum Driving Force (MDF) | max min(–ΔG'i) across all steps [88] | >0 kJ/mol | Identifies thermodynamic bottlenecks; higher values enable higher fluxes |
| Flux-Force Efficacy (FFE) | (e^(ΔGdiss/RT) – 1)/(e^(ΔGdiss/RT) + 1) [88] | 0.8–1.0 | Quantifies enzyme utilization efficiency; values near 1 indicate minimal reverse flux |
| Energy Yield (ATP) | ATP produced per substrate molecule [88] | Pathway-dependent | Maximized in energy-limited environments; trade-off with rate |
| NADPH Regeneration Rate | mmol NADPH/gDCW/h [48] | Matches pathway demand | Critical for anabolic pathways; balanced with carbon flux |
Table 2: Successful Cofactor Engineering Strategies in Microbial Factories
| Target Molecule | Host Organism | Key Cofactor Engineering Strategy | Yield Improvement | Reference |
|---|---|---|---|---|
| D-pantothenic acid | E. coli | Integrated transhydrogenase + TCA flux control + ATP synthase tuning [48] | 86.03 g/L in 5L fermenter [48] | [48] |
| Riboflavin | E. coli | PPP flux enhancement + purine biosynthesis deregulation + NADPH/ATP balancing [87] | Not specified | [87] |
| 1,4-butanediol | E. coli | Multiplex experimentation + machine learning-guided optimization [89] | Commercial scale achieved [89] | [89] |
Experimental Protocols for Pathway Optimization
Protocol 3: Multiplex Pathway Variant Construction and Screening
Principle: Simultaneously test multiple enzyme combinations and regulatory elements using pooled libraries to rapidly identify optimal pathway configurations [89].
Procedure:
- Library Design: Design a combinatorial library of pathway variants with different promoters, RBS sequences, and enzyme orthologs using automated DNA assembly.
- Pooled Transformation: Transform the entire library into the host organism simultaneously, creating a vast array of pathway variants in a single experiment.
- High-Throughput Screening: Use microdroplet-assisted screening or robotic systems to isolate high-performing variants based on product-specific fluorescence reporters or growth coupling [89].
- Machine Learning Analysis: Apply random forest or neural network models to extract design rules from screening data, identifying synergistic combinations for further optimization [89].
Protocol 4: Driving Force Optimization for Thermodynamic Feasibility
Principle: Adjust metabolite concentrations to maximize the minimum driving force across the pathway, improving both thermodynamic feasibility and kinetic efficiency [88].
Procedure:
- Thermodynamic Data Collection: Compile standard Gibbs free energy values (ΔG'°) for all pathway reactions from databases or group contribution methods.
- Concentration Bounds Definition: Set physiologically relevant concentration ranges for all metabolites (typically 0.001–10 mM).
- Optimization Problem Formulation: Formulate as a multi-objective mixed-integer linear programming problem maximizing both MDF and ATP yield [88].
- Concentration Optimization: Solve using ε-constraint method to find metabolite concentrations that maximize MDF while maintaining pathway feasibility [88].
Pathway Analysis and Visualization
Conceptual Workflow for Pathway Variant Analysis
Integrated Cofactor Regulation Network
Research Reagent Solutions
Table 3: Essential Research Reagents for Cofactor Engineering Studies
| Reagent/Category | Specific Examples | Research Application | Key Function in Pathway Engineering |
|---|---|---|---|
| Enzyme Expression | pET vectors, pRSF vectors [89] | Heterologous pathway expression | Tunable protein production for balancing metabolic flux |
| Cofactor Regeneration | Zwf, PntAB, GapN [48] | NADPH enhancement systems | Increase reducing equivalent supply for anabolic reactions |
| Flux Analysis Tools | FBA, FVA, MDF optimization [88] [48] | Metabolic network modeling | Predict flux distributions and identify thermodynamic bottlenecks |
| Genome Editing | CRISPR-Cas9, MAGE [89] | Multiplex strain engineering | Enable simultaneous modification of multiple genomic targets |
| Analytical Standards | NADPH, ATP, pathway intermediates [48] | Metabolite quantification | Accurate measurement of intracellular metabolite concentrations |
Compartment-Specific Cofactor Ratio Measurements Using Sensor Reactions
Troubleshooting Guides
Common Sensor Issues and Solutions
Table 1: Troubleshooting NAD(P)H Sensor Problems
| Problem Category | Specific Issue | Possible Causes | Recommended Solutions |
|---|---|---|---|
| Signal Quality | Prolonged response time [90] | Sensor aging, suboptimal temperature | Confirm operating environment is within sensor specifications [90] |
| Reduced accuracy [90] | pH interference, photobleaching | Use cpYFP control to normalize for pH changes [91]; check calibration | |
| Signal instability/zero drift [90] | Temperature fluctuations, power supply voltage instability, component aging | Stabilize environmental conditions; verify power supply; replace aged sensor [90] | |
| Reduced sensitivity [90] | Sensor overload, general performance degradation | Avoid inputs exceeding design specifications; replace damaged sensor [90] | |
| Technical Failures | Overload damage [90] | Input signal exceeds sensor's maximum range | Replace sensor; ensure input signals are within specified range [90] |
| Electrical failure [90] | Short circuits, broken wires, poor contact | Perform visual inspection of wires/connectors; use multimeter to test circuits [90] | |
| Signal interference [90] | External electromagnetic noise | Identify/remove EMI sources; use shielding and isolation measures [90] | |
| Experimental Validity | Unexpected ratio readings | Compartmental specificity loss, incorrect targeting | Verify targeting with fluorescent markers (e.g., TMRM for mitochondria) [91] |
| Poor response to metabolic perturbations | Incorrect permeabilization, non-physiological conditions | Optimize digitonin concentration for membrane permeabilization [91] |
Frequently Asked Questions (FAQs)
Q1: Why is it crucial to measure NAD+/NADH and NADPH/NADP+ ratios in specific cellular compartments rather than just whole-cell levels? A: Subcellular compartments maintain distinct and separate NAD(P)H pools. The mitochondrial pool is separated from the cytosolic pool because the inner mitochondrial membrane is impermeable to NAD/NADH [91]. These pools respond differently to metabolic perturbations and pathophysiological conditions. Bulk cellular measurements homogenize these distinct pools and can mask critical, compartment-specific redox dynamics that are essential for understanding cellular function and engineering metabolic pathways [91] [92].
Q2: What is the advantage of using genetically encoded biosensors like SoNar over traditional methods like enzymatic cycling assays or NADH autofluorescence? A: Genetically encoded biosensors provide real-time, compartment-specific data in live cells, unlike endpoint enzymatic assays that require homogenization [91]. SoNar also directly reports the NAD+/NADH ratio, whereas NADH autofluorescence cannot distinguish NADH from NADPH and does not provide ratio information, leading to potential misinterpretation [91]. Additionally, biosensors targeted to specific locations (e.g., mt-SoNar for mitochondria) offer superior spatial resolution and specificity [91].
Q3: My sensor readings are fluctuating unexpectedly. How can I determine if this is due to a real change in the NAD+/NADH ratio or an artifact from pH changes? A: The cpYFP component of sensors like SoNar is pH-sensitive. The standard solution is to co-express a pH-control sensor such as mt-cpYFP (for mitochondria) or ct-cpYFP (for cytosol). The NAD+/NADH ratio should be derived from the SoNar fluorescence signal after normalization to the signal from the pH-control sensor, which effectively eliminates the interference from potential pH changes [91].
Q4: In the context of engineered pathways, what are the consequences of an imbalanced NADPH/NADP+ ratio? A: Redox imbalance can halt metabolic flux, waste carbon and energy, and compromise cell viability [58] [23]. In production strains, a lack of NADPH can be a limiting factor for yield, as seen in L-threonine biosynthesis [58]. Conversely, strategic creation of a redox imbalance driving force (RIFD) by generating excess NADPH can be used to direct carbon flux toward a desired product, restoring growth and enhancing production [58].
Q5: How can I validate that a sensor targeted to a specific compartment, like the mitochondria, is functioning correctly and responding to known metabolic perturbations? A: You can use well-established metabolic modulators to challenge the system. For a mitochondrial sensor (mt-SoNar), applying 1 mM β-hydroxybutyrate (β-OHB) should decrease the NAD+/NADH ratio, while 1 mM acetoacetic acid (AcAc) should increase it, based on the equilibrium of the mitochondrial enzyme β-hydroxybutyrate dehydrogenase [91]. A correct response to these compounds confirms the sensor is functioning appropriately in its intended compartment.
Experimental Protocols
Protocol 1: Monitoring Compartmentalized NAD+/NADH Ratios with Targeted SoNar
Objective: To measure real-time changes in the NAD+/NADH ratio within the cytosol and mitochondria of live cells using the genetically encoded biosensor SoNar [91].
Key Reagents:
- mt-SoNar (SoNar targeted to the mitochondrial matrix via 2xCOXVIII targeting sequence)
- ct-SoNar (cytosolic-targeted SoNar)
- mt-cpYFP or ct-cpYFP (pH control sensors)
- Appropriate cell line (e.g., H9C2 cardiomyoblasts, adult rat cardiomyocytes)
- TMRM (mitochondrial membrane potential dye for localization validation)
- Digitonin
- Metabolic modulators: β-hydroxybutyrate (β-OHB), Acetoacetic acid (AcAc)
Methodology:
- Sensor Expression: Transduce cells with lentiviral constructs for mt-SoNar/ct-SoNar and the corresponding mt-cpYFP/ct-cpYFP.
- Localization Validation: Confirm mitochondrial localization by co-staining with TMRM and performing confocal microscopy [91].
- In Situ Calibration (Optional): Permeabilize the plasma membrane (0.001% digitonin) or mitochondrial membrane (0.3% digitonin) and expose cells to buffers with defined NAD+/NADH ratios (from 1000:1 to 1:1) to establish a linear response curve [91].
- Live-Cell Imaging: Conduct ratiometric imaging (excitation at 410 nm and 480 nm, emission at 520 nm) using a confocal microscope.
- Data Analysis: Calculate the ratio of SoNar fluorescence (F480/F410). Normalize this ratio to the signal from cpYFP to correct for pH artifacts. The final output reflects the NAD+/NADH ratio [91].
- Metabolic Challenge: Apply interventions like 1 mM β-OHB or 1 mM AcAc, or simulate ischemia/reperfusion to observe dynamic ratio changes [91].
Protocol 2: Tracing Compartmentalized NADPH Metabolism Using Stable Isotopes
Objective: To resolve the contributions of different pathways (e.g., pentose phosphate pathway, mitochondrial one-carbon metabolism) to cytosolic and mitochondrial NADPH pools [92].
Key Reagents:
- 2H-labeled substrates (e.g., [2H] glucose)
- Cells expressing compartment-specific reporters (e.g., cytosolic or mitochondrial IDH mutants)
- LC-MS (Liquid Chromatography-Mass Spectrometry) system
Methodology:
- Reporter System: Utilize cells expressing enzymes that produce a trackable metabolite in a specific compartment, such as mutant isocitrate dehydrogenase (IDH) which produces 2-hydroxyglutarate (2HG) using NADPH [92].
- Isotope Labeling: Feed cells 2H-labeled substrates. The hydrogen from NADPH is incorporated into the product (e.g., 2HG).
- Compartment-Specific Tracing: Measure the 2H-labeling pattern in 2HG produced by cytosolic vs. mitochondrial targeted reporters. This reports on the NADPH used in each location.
- Pathway Inhibition: Use specific inhibitors (e.g., of the PPP) to quantify how much different pathways contribute to NADPH production in each compartment [93].
- MS Analysis: Extract metabolites and analyze labeling patterns via LC-MS to determine the origin of hydrogens in NADPH-dependent products [92].
Research Reagent Solutions
Table 2: Essential Research Reagents for Cofactor Sensing
| Reagent Name | Function/Application | Key Characteristics |
|---|---|---|
| SoNar Biosensor | Ratiometric monitoring of NAD+/NADH | Genetically encoded; two excitation peaks (410/480 nm); one emission peak (520 nm); can be targeted to organelles [91] |
| LbNOX / mitoLbNOX | Genetically manipulating NAD+/NADH ratio | Enzyme from L. brevis; consumes NADH to increase NAD+/NADH ratio; mitoLbNOX is mitochondria-targeted [94] |
| Cytosolic Malic Enzyme (ME1) | Provides cytosolic NADPH | Catalyzes malate to pyruvate conversion; generates NADPH; rescues redox defects in CI-deficient cells [93] |
| PdxJ / YqhD | NADPH-consuming / producing modules | Used in metabolic engineering to create synthetic NADPH sinks or sources for redox balancing [58] |
| 2H-Labeled Substrates | Tracing NADPH metabolism | Used with LC-MS to track hydrogen from NADPH into specific products, resolving compartmentalized pathway activity [92] |
| Nicotinamide Nucleotide Transhydrogenase (NNT) | Mitochondrial NADPH generation | Inner mitochondrial membrane enzyme; transfers hydride from NADH to NADP+; key for mitochondrial NADPH supply [95] |
Signaling Pathways and Workflows
Diagram Title: Cofactor Sensing Workflow and Pitfalls
Diagram Title: Compartmentalized NADPH Metabolism Pathways
In the realm of bioprocessing, maintaining precise cofactor balance is not merely an optimization strategy but a fundamental requirement for robust and validated production systems. Nicotinamide adenine dinucleotide phosphate (NADPH) and adenosine triphosphate (ATP) are pivotal cofactors, acting as the primary currency of reducing power and energy, respectively. Efficient microbial factories and compliant biomedical production systems depend on their careful management. Insufficient NADPH supply can lead to cell death under reactive oxygen species (ROS) and limits the production of high-value chemicals, while ATP availability is extremely important for cell growth and biosynthesis [96] [27]. This technical support center provides targeted troubleshooting guides and FAQs, framed within a broader thesis on managing cofactor balance, to help researchers address specific challenges in their experiments.
Cofactor Engineering Fundamentals: Key Concepts for Troubleshooting
The Metabolic Pathways for NADPH and ATP Regeneration
Understanding the core metabolic pathways is the first step in effective troubleshooting. The following diagram illustrates the primary routes for NADPH and ATP generation and their engineering strategies.
The efficient regeneration of NADPH and ATP is a determinant of cellular energy availability and a common bottleneck in production [96] [27]. The oxidative pentose phosphate pathway (oxPPP) is the main inherent route for NADPH generation in many microorganisms, catalyzed by key enzymes like glucose-6-phosphate dehydrogenase (ZWF1/G6PDH) and 6-phosphogluconate dehydrogenase (GND2/6PGDH) [96] [27]. However, static over-expression of these pathways often leads to an imbalance in the NADPH/NADP+ ratio, causing disruptions in cell growth and production [27]. Dynamic regulation strategies, including the use of genetically encoded biosensors, are emerging as superior solutions for real-time monitoring and control of the intracellular NADP(H) redox status [27].
The Scientist's Toolkit: Essential Research Reagent Solutions
The table below details key reagents, strains, and genetic tools used in cofactor engineering experiments, along with their specific functions.
Table 1: Key Research Reagents and Tools for Cofactor Engineering
| Item Name | Type/Function | Example Application in Research |
|---|---|---|
| ZWF1/gcdA Gene | Encodes Glucose-6-phosphate Dehydrogenase (G6PDH); key enzyme in oxPPP [96] [47]. | Overexpression in Pichia pastoris and Aspergillus niger to increase NADPH supply for terpene and protein production [96] [47]. |
| GND2/gndA Gene | Encodes 6-Phosphogluconate Dehydrogenase (6PGDH); a major NADPH source in oxPPP [96] [47]. | Overexpression in A. niger led to a nine-fold increase in intracellular NADPH concentration and a 65% increase in glucoamylase yield [47]. |
| cPOS5/POS5 Gene | Encodes a NADH kinase; converts NADH to NADPH, creating an alternative regeneration route [96]. | Heterologous expression from S. cerevisiae in P. pastoris at low intensity to aid α-farnesene production without overly disrupting redox balance [96]. |
| maeA Gene | Encodes a NADP-dependent malic enzyme; generates NADPH outside the PPP [47]. | Overexpression in A. niger increased the intracellular NADPH pool by 66% and glucoamylase yield by 30% [47]. |
| SoxR & NERNST | Genetically encoded biosensors for detecting intracellular NADPH/NADP+ ratio [27]. | Enable real-time monitoring and dynamic regulation of the NADP(H) redox status in live cells [27]. |
| Tet-On Gene Switch | Tight, tunable, and metabolism-independent promoter system for controlled gene expression [47]. | Used in A. niger to precisely overexpress NADPH-generating genes and study their individual effects [47]. |
| APRT Gene | Encodes Adenine Phosphoribosyltransferase; involved in AMP/ATP biosynthesis [96]. | Overexpression in P. pastoris to increase the supply of adenosine monophosphate (AMP) for ATP synthesis [96]. |
| GPD1 Gene | Encodes Glycerol-3-phosphate dehydrogenase; a competing pathway for NADH consumption [96]. | Inactivation in P. pastoris to conserve NADH, which is crucial for ATP regeneration via electron transport phosphorylation [96]. |
Troubleshooting Guides and FAQs: Addressing Specific Experimental Issues
This section provides direct, actionable answers to common problems encountered during experiments related to cofactor balance and bioprocess validation.
FAQ: Microbial Production and Cofactor Engineering
Q1: My microbial production strain for a NADPH-intensive product (e.g., a terpene) is underperforming despite a engineered pathway. The strain also shows poor growth. What could be the issue?
A: This is a classic symptom of cofactor imbalance. Your engineering strategy may have created a static over-demand for NADPH that the host's native metabolism cannot meet, hampering both growth and production [27].
- Troubleshooting Steps:
- Quantify Cofactor Levels: If possible, measure the intracellular NADPH/NADP+ ratio. A severely low ratio confirms insufficient regeneration capacity [27].
- Avoid Simple Overexpression: Instead of merely overexpressing a single PPP gene like ZWF1, consider a combined approach. In P. pastoris, simultaneously overexpressing ZWF1 and SOL3 was more effective than ZWF1 alone for α-farnesene production [96].
- Explore Alternative NADPH Sources: Introduce or overexpress genes from other NADPH-generating pathways. The expression of maeA (NADP-dependent malic enzyme) or gndA (6-phosphogluconate dehydrogenase) has successfully boosted production in A. niger [47].
- Implement Dynamic Regulation: For a more advanced solution, develop a dynamic control system using an NADPH-sensitive biosensor (e.g., SoxR). This can dynamically regulate pathway genes in response to real-time cofactor demand, avoiding growth defects [27].
Q2: I have engineered the NADPH supply successfully, but my product yield is still not meeting theoretical expectations. What other cofactor should I investigate?
A: It is crucial to remember that ATP is often a co-requisite with NADPH in many biosynthesis pathways. For example, the production of one molecule of α-farnesene via the mevalonate pathway requires 6 molecules of NADPH and 9 molecules of ATP [96]. An ATP bottleneck can stall your process.
- Troubleshooting Steps:
- Engineer ATP Regeneration: Focus on strategies to enhance ATP availability. In P. pastoris, this was achieved by overexpressing APRT to increase the AMP supply and inactivating GPD1 to reduce wasteful NADH consumption, thereby making more NADH available for ATP synthesis via oxidative phosphorylation [96].
- Check for Energy Spills: Investigate if there are any ATP-consuming futile cycles or stress responses that are unnecessarily depleting the ATP pool.
Q3: What is a robust experimental protocol for testing the impact of different NADPH-enhancing genes in a new host strain?
A: A systematic protocol, inspired by the Design-Build-Test-Learn (DBTL) cycle used in Aspergillus niger, is recommended [47].
- Detailed Methodology:
- Design:
- Select a panel of candidate genes from different pathways (e.g., zwf1/gcdA from PPP, maeA from TCA, cPOS5 as a kinase).
- Choose a consistent, neutral genomic integration site (e.g., pyrG locus) to avoid position effects.
- Use a uniform, inducible promoter system (e.g., the Tet-On switch) for all constructs to enable fair comparison and control expression levels [47].
- Build:
- Use CRISPR/Cas9 or other precise genetic tools to integrate each candidate gene into your production host, creating a set of isogenic strains differing only in the engineered NADPH module.
- Test:
- Cultivate all strains in shake flasks under controlled, reproducible conditions.
- Measure key performance indicators (KPIs): product titer, total protein, biomass yield.
- For the most promising strains, perform chemostat cultivations for more precise metabolomic analysis. Quantify intracellular NADPH/NADP+ ratios and relevant metabolic fluxes [47].
- Learn:
- Analyze which genetic modifications provided the most significant boost to your specific product.
- Use this data to inform the next DBTL cycle, potentially testing combinations of the most effective genes.
- Design:
FAQ: Biomedical Production and Validation
Q4: During the manufacture of a biologic drug substance, we are consistently seeing bioburden excursions in our in-process pools. Our risk assessment deemed them "low-risk," but a regulatory inspection cited us. What went wrong?
A: The deficiency likely stemmed from an inadequate microbial risk assessment and process design. Relying on a flawed methodology, such as converting colony-forming units (CFU) to a "mass-based exposure" to justify out-of-limit batches, is not acceptable to regulators. Contaminants can introduce impurities, endotoxins, and cause product degradation, impacting quality beyond a simple mass calculation [97] [98].
- Corrective Actions:
- Strengthen Your Control Strategy: The process must be designed to prevent contamination, not just to filter it out later. Contamination should be strictly avoided with appropriate in-process action limits [97] [98].
- Conduct a Root Cause Investigation: Systematically examine all potential entry points for microbial contamination. Common issues include:
- Damaged or missing O-rings on probes and valves.
- Improperly installed or deformed air filters after sterilization.
- Incorrectly sloped condensate lines leading to back-flow.
- Problems with sampling devices and addition valves [98].
- Enhance Preventative Maintenance: Implement a rigorous preventative maintenance plan for all fermentor/bioreactor components, including valves, diaphragms, gaskets, and filters [98].
- Validate Sterility: Perform media simulations to demonstrate the sterility of your equipment and processes, especially after shutdowns or maintenance [98].
Q5: In our aseptic filling suite, investigators observed operators blocking "first air" on sterile containers and adjusting filling needles with non-sterile gloves. Why are these practices major deficiencies?
A: These observations directly violate the core principles of aseptic processing. The "first air" from HEPA filters is the sterile air that provides a protective blanket over critical zones. Blocking it with hands, arms, or equipment introduces unsterile airflow and particulate matter. Similarly, touching critical sterile path components (like filling needles) with non-sterile gloves is a direct route for microbial contamination [97].
- Corrective Actions:
- Re-train Personnel: All staff must be thoroughly trained and periodically evaluated on aseptic techniques, including moving slowly and deliberately and understanding the concept of "first air" protection [97].
- Establish and Follow Strict SOPs: Procedures must explicitly forbid blocking first air and handling critical sterile components with anything but sterile tools or gloved hands that have been properly sanitized [97].
- Review Gowning Procedures: Ensure that personnel are wearing appropriate sterile gowning and gloves.
- Review and Sterilize RABS Gloves: Restrictive Access Barrier System (RABS) gloves should be periodically sterilized and not reused for extended periods (e.g., 100 batches) without sterilization [97].
Comparative Analysis: Quantitative Data from Cofactor Engineering Case Studies
The table below summarizes key performance data from published case studies where cofactor engineering was applied, providing a benchmark for expected outcomes.
Table 2: Quantitative Outcomes from Cofactor Engineering Case Studies
| Host Organism | Target Product | Engineering Strategy | Key Performance Outcome | Citation |
|---|---|---|---|---|
| Pichia pastoris | α-Farnesene | Combined overexpression of ZWF1 & SOL3 (oxPPP). | Increased α-farnesene production by ~8.7% and ~12.9%, respectively. | [96] |
| Pichia pastoris | α-Farnesene | Low-intensity expression of cPOS5 (NADH kinase) + APRT overexpression & GPD1 inactivation (ATP). | Final strain P. pastoris X33-38 produced 3.09 ± 0.37 g/L, a 41.7% increase over the parent strain. | [96] |
| Aspergillus niger | Glucoamylase (GlaA) | Overexpression of gndA gene (6-phosphogluconate dehydrogenase). | Increased intracellular NADPH pool by 45% and GlaA yield by 65% in chemostat cultures. | [47] |
| Aspergillus niger | Glucoamylase (GlaA) | Overexpression of maeA gene (NADP-dependent malic enzyme). | Increased intracellular NADPH pool by 66% and GlaA yield by 30% in chemostat cultures. | [47] |
| E. coli | General NADPH Supply | Introduction of synthetic Entner–Doudoroff pathway from Zymomonas mobilis. | Increased the NADPH regeneration rate 25-fold. | [96] |
Successful bioproduction, from the microbial factory to the cGMP suite, requires an integrated view of intracellular metabolism and external production controls. As the case studies demonstrate, rationally modifying NADPH and ATP regeneration pathways can dramatically improve titers and yields [96] [47]. However, these gains are only sustainable in a regulated environment when coupled with a robust microbial control strategy that proactively prevents contamination through rigorous equipment design, preventative maintenance, and strict adherence to aseptic procedures [97] [98]. By applying the troubleshooting guides and foundational knowledge presented here, researchers and drug development professionals can more effectively design experiments, diagnose failures, and develop validated, industrial-strength production systems.
Conclusion
Effective management of NADPH and ATP cofactor balance represents a cornerstone of successful metabolic engineering, transcending traditional pathway optimization to address fundamental cellular energy and redox economics. The integration of computational modeling with advanced engineering strategies enables precise control over cofactor metabolism, leading to significantly enhanced production yields in biomedical and industrial applications. Future directions will likely focus on dynamic regulation systems, orthogonal cofactor engineering, and the development of cross-kingdom solutions that leverage photosynthetic organisms for sustainable cofactor regeneration. As genetically encoded biosensors and multi-omics technologies advance, real-time monitoring and adjustment of cofactor balances will become increasingly sophisticated, opening new frontiers in therapeutic compound production, personalized medicine, and sustainable biomedicine. The continued convergence of synthetic biology, systems biology, and metabolic engineering promises to transform cofactor management from a persistent challenge into a powerful design feature for next-generation bioproduction platforms.
References
- 1. NADPH—The Forgotten Reducing Equivalent - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC8168431/]
- 2. Reduced nicotinamide adenine dinucleotide phosphate in redox balance and diseases: a friend or foe? - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC9343382/]
- 3. for Efficient Production of α-Farnesene by... Cofactor Engineering [https://www.mdpi.com/1422-0067/24/2/1767]
- 4. and ATP engineering of Escherichia coli to improve the... NADPH [https://biotechnologyforbiofuels.biomedcentral.com/articles/10.1186/s13068-021-01954-6]
- 5. Mitochondrial complex I inhibition triggers NAD+-independent glucose... [https://link.springer.com/article/10.1007/s11357-023-01059-y]
- 6. Different Biochemical Mechanisms Ensure Network-Wide Balancing of... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2655529/]
- 7. NADPH homeostasis in cancer: functions, mechanisms and therapeutic implications | Signal Transduction and Targeted Therapy [https://www.nature.com/articles/s41392-020-00326-0]
- 8. Redox Engineering for Improved... Cofactor Balance [https://pubmed.ncbi.nlm.nih.gov/38625685/]
- 9. In silico model-driven cofactor strategies for improving... engineering [https://link.springer.com/article/10.1007/s10295-015-1663-0]
- 10. Acetol biosynthesis enables NADPH during nitrogen limitation... balance [https://microbialcellfactories.biomedcentral.com/articles/10.1186/s12934-025-02687-z]
- 11. Overview of Pentose Phosphate Pathway - Creative Proteomics [https://metabolomics.creative-proteomics.com/resource/overview-pentose-phosphate-pathway.htm]
- 12. ATP/ADP [https://chem.libretexts.org/Bookshelves/Biological_Chemistry/Supplemental_Modules_(Biological_Chemistry)/Metabolism/ATP_ADP]
- 13. 20.1 ATP—the Universal Energy Currency [https://courses.lumenlearning.com/suny-orgbiochemistry/chapter/20-1-atp-the-universal-energy-currency/]
- 14. 20.2: ATP- the Universal Energy Currency [https://chem.libretexts.org/Courses/Matanuska-Susitna_College/MatSu_College-CHEM_A104_Introduction_to_Organic_and_Biochemistry/20%3A_Energy_Metabolism/20.02%3A_ATP-_the_Universal_Energy_Currency]
- 15. ATP- The universal energy currency of cell | PPT [https://www.slideshare.net/slideshow/atp-the-universal-energy-currency-of-cell/57407266]
- 16. In silico co-factor balance estimation using constraint-based ... [https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1008125]
- 17. 10 Common ATP Test Failures (And How to Fix High RLU ... [https://empacsgroup.com/10-common-atp-test-failures-and-how-to-fix-high-rlu-scores/?srsltid=AfmBOoqYdYez_Y1JXJcWJHuHUdc3Sgmk0NXABjRtfqLDtaSIcAf7ymix]
- 18. A general tool for engineering the NAD/NADP cofactor ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8611728/]
- 19. Engineering cofactor metabolism for improved protein and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7584080/]
- 20. Metabolic Impact of Redox Cofactor Perturbations on the ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4702612/]
- 21. Principles and functions of metabolic compartmentalization [https://pmc.ncbi.nlm.nih.gov/articles/PMC10155461/]
- 22. Evaluating the contribution of plant metabolic pathways in the ... [https://pubmed.ncbi.nlm.nih.gov/38874662/]
- 23. Review Engineering redox balance through cofactor systems [https://www.sciencedirect.com/science/article/abs/pii/S0167779914000675]
- 24. Engineering Compartmentalization and Cofactor ... [https://pubmed.ncbi.nlm.nih.gov/40354551/]
- 25. Defining the ATPome reveals cross-optimization of ... [https://www.nature.com/articles/s41467-020-18084-6]
- 26. Metabolic Engineering Problem [https://neos-guide.org/case-studies/bam/metabolic-engineering-problem/]
- 27. Dynamic and Static Regulation of Nicotinamide Adenine ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11314019/]
- 28. A versatile in situ cofactor enhancing system for meeting ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10850783/]
- 29. Optimal cofactor swapping can increase the theoretical ... [https://www.sciencedirect.com/science/article/abs/pii/S1096717614000688]
- 30. Multiple Cofactor Engineering Strategies to Enhance ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11123869/]
- 31. Multiple Cofactor Engineering Strategies to Enhance ... [https://www.mdpi.com/2076-2607/12/5/933]
- 32. Cofactor engineering in cyanobacteria to overcome ... [https://link.springer.com/article/10.1007/s11705-016-1591-1]
- 33. In silico co-factor balance estimation using constraint-based ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7440669/]
- 34. In silico co-factor balance estimation using constraint ... [https://journals.plos.org/ploscompbiol/article/figures?id=10.1371/journal.pcbi.1008125]
- 35. Quantifying the propagation of parametric uncertainty on ... [https://www.sciencedirect.com/science/article/pii/S1096717621001634]
- 36. Genetically encoded ATP and NAD(P)H biosensors [https://pubmed.ncbi.nlm.nih.gov/36130803/]
- 37. Cofactor engineering - Wikipedia [https://en.wikipedia.org/wiki/Cofactor_engineering]
- 38. Frontiers | Protein Engineering for Nicotinamide Coenzyme Specificity in Oxidoreductases: Attempts and Challenges [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2018.00194/full]
- 39. Elucidation of new condition-dependent roles for... | PLOS One [https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0177319]
- 40. A synthetic biochemistry molecular purge valve module ... [https://www.nature.com/articles/ncomms5113]
- 41. A synthetic biochemistry molecular purge valve module ... [https://pubmed.ncbi.nlm.nih.gov/24936528/]
- 42. A Hitchhiker's Guide to Supplying Enzymatic Reducing Power ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10127272/]
- 43. A synthetic biochemistry molecular purge valve module that ... [https://patents.google.com/patent/WO2015153929A3/en]
- 44. Research team led by Professor James Bowie develops “ ... [https://www.chemistry.ucla.edu/news/research-team-led-professor-james-bowie-develops-synthetic-biochemistry-bypasses-need-cells/]
- 45. New tools for redox biology: From imaging to manipulation [https://www.sciencedirect.com/science/article/abs/pii/S0891584916310929]
- 46. Protocol for building synthetic protocell membranes that ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11264183/]
- 47. Engineering cofactor metabolism for improved protein and ... [https://microbialcellfactories.biomedcentral.com/articles/10.1186/s12934-020-01450-w]
- 48. Integrated cofactor-centric engineering and energy flux ... [https://www.sciencedirect.com/science/article/abs/pii/S1385894725100223]
- 49. Cofactor Engineering for Enhancing the Flux of Metabolic ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4147997/]
- 50. Pathway design using de novo steps through uncharted ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5766603/]
- 51. Cofactor engineering for more efficient production of chemicals and biofuels - ScienceDirect [https://www.sciencedirect.com/science/article/abs/pii/S0734975017301192]
- 52. Frontiers | Application of cofactors in the regulation of microbial metabolism: A state of the art review [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2023.1145784/full]
- 53. The Importance of Energy Balance in Improving Photosynthetic Productivity - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC3075755/]
- 54. The challenges of scaling biofuel production. [https://www.rentechinc.com/2025/10/09/the-challenges-of-scaling-biofuel-production/]
- 55. Environmental analysis of biotechnologies for biofuels , bioplastics ... [https://biotechforenvironment.biomedcentral.com/articles/10.1186/s44314-024-00010-5]
- 56. Biobased production of alkanes and alkenes through metabolic... [https://link.springer.com/article/10.1007/s10295-016-1814-y]
- 57. Genome-Scale Consequences of Cofactor Balancing in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3208632/]
- 58. Developing a Redox Imbalance Forces Drive (RIFD) ... [https://www.sciencedirect.com/science/article/abs/pii/S1096717625001181]
- 59. Review Self-assembly systems to troubleshoot metabolic ... [https://www.sciencedirect.com/science/article/abs/pii/S0167779923001877]
- 60. A versatile in situ cofactor enhancing system for meeting ... [https://www.sciencedirect.com/science/article/pii/S0021925823026273]
- 61. Evaluating Cell Metabolism Through Autofluorescence ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6352511/]
- 62. Metabolic profiling of single cells by exploiting NADH and ... [https://www.sciencedirect.com/science/article/pii/S2212877824001121]
- 63. Futile cycle [https://en.wikipedia.org/wiki/Futile_cycle]
- 64. Acetol biosynthesis enables NADPH balance during nitrogen ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11910842/]
- 65. Physiology, Adenosine Triphosphate - StatPearls - NCBI - NIH [https://www.ncbi.nlm.nih.gov/books/NBK553175/]
- 66. Mitochondria-derived nuclear ATP surge protects against ... [https://www.nature.com/articles/s41467-025-61787-x]
- 67. Construction of an alternative NADPH regeneration ... [https://www.sciencedirect.com/science/article/pii/S2405805X24000292]
- 68. Engineering Yarrowia lipolytica for the production of β ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11734496/]
- 69. NADPH-dependent and -independent Disulfide Reductase ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6165701/]
- 70. Rerouting of NADPH synthetic pathways for increased ... [https://www.nature.com/articles/s41598-018-34210-3]
- 71. Activation of the pentose phosphate pathway by ... [https://www.frontiersin.org/journals/physiology/articles/10.3389/fphys.2025.1666999/full]
- 72. Enforced ATP Futile Cycling Increases Specific Productivity ... [https://pubmed.ncbi.nlm.nih.gov/25899755/]
- 73. Cofactor engineering for advancing chemical biotechnology [https://www.sciencedirect.com/science/article/abs/pii/S0958166913000797]
- 74. ATP-consuming futile cycles as energy dissipating ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8873062/]
- 75. Designing pathways for bioproducing complex chemicals ... [https://www.nature.com/articles/s41467-025-59827-7]
- 76. Heterologous Metabolic Pathways: Strategies for Optimal ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7385092/]
- 77. Application of cofactors in the regulation of microbial metabolism [https://pmc.ncbi.nlm.nih.gov/articles/PMC10126289/]
- 78. Metabolic reconfiguration enables synthetic reductive ... [https://www.nature.com/articles/s42255-022-00654-1]
- 79. A family of NADPH/NADP+ biosensors reveals in vivo ... [https://www.nature.com/articles/s41467-024-55302-x]
- 80. A family of NADPH/NADP+ biosensors reveals in vivo ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11659435/]
- 81. Methods for Using a Genetically Encoded Fluorescent ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6378224/]
- 82. Guidelines on experimental methods to assess ... [https://www.nature.com/articles/s41418-017-0020-4]
- 83. Genetically encoded tools for measuring and manipulating ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9093021/]
- 84. What is flux balance analysis? - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC3108565/]
- 85. Flux balance analysis [https://en.wikipedia.org/wiki/Flux_balance_analysis]
- 86. Flux balance analysis accounting for metabolite dilution [https://genomebiology.biomedcentral.com/articles/10.1186/gb-2010-11-4-r43]
- 87. Recent advances in metabolic engineering of Escherichia ... [https://pubmed.ncbi.nlm.nih.gov/40999135/]
- 88. Optimal evaluation of energy yield and driving force in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10353829/]
- 89. Enabling pathway design by multiplex experimentation and ... [https://www.sciencedirect.com/science/article/abs/pii/S1096717623001702]
- 90. Common Sensor Troubleshooting Methods - WFsensors [https://www.wfsensors.com/blog/common-sensor-troubleshooting-methods.html/]
- 91. Genetically encoded biosensors for evaluating NAD+ ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8659198/]
- 92. Tracing Compartmentalized NADPH Metabolism in the ... [https://www.sciencedirect.com/science/article/pii/S109727651400402X]
- 93. Defective NADPH production in mitochondrial disease ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7264245/]
- 94. The compartment-specific manipulation of the NAD+/NADH ... [https://www.nature.com/articles/s41598-024-71462-8]
- 95. Cofactor Balance by Nicotinamide Nucleotide ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3642339/]
- 96. Cofactor Engineering for Efficient Production of α-Farnesene ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9860691/]
- 97. Biologics Inspection Case Studies Highlight API and Drug ... [https://redica.com/biologics-inspection-case-studies-highlight-api-and-drug-product-deficiencies/]
- 98. Case Studies of Microbial Contamination in Biologic ... [https://www.americanpharmaceuticalreview.com/Featured-Articles/36755-Case-Studies-of-Microbial-Contamination-in-Biologic-Product-Manufacturing/]