Dynamic Regulation of NADPH/NADP+ Balance: A Biosensor-Driven Approach for Metabolic Engineering and Drug Discovery
The NADPH/NADP+ redox couple is a central regulator of cellular redox homeostasis, antioxidant defense, and reductive biosynthesis.
Dynamic Regulation of NADPH/NADP+ Balance: A Biosensor-Driven Approach for Metabolic Engineering and Drug Discovery
Abstract
The NADPH/NADP+ redox couple is a central regulator of cellular redox homeostasis, antioxidant defense, and reductive biosynthesis. Imbalances in this ratio can disrupt cell growth and limit the production of high-value biochemicals and therapeutics. This article explores the revolutionary role of genetically encoded biosensors in overcoming these challenges. We cover the foundational principles of NADP(H) metabolism, detail the design and real-world application of cutting-edge biosensors like the NAPstar family, and provide troubleshooting strategies for implementation. A comparative analysis validates the performance of various biosensor platforms. Aimed at researchers and drug development professionals, this review synthesizes current knowledge to provide a roadmap for using dynamic biosensor-driven regulation to advance metabolic engineering and biomedical research.
The Critical Role of NADPH/NADP+ Balance in Cellular Redox and Why It Matters
Nicotinamide adenine dinucleotide phosphate (NADPH) serves as an essential electron donor across all domains of life, functioning as the cell's primary reducing power for both antioxidant defense and reductive biosynthesis [1]. The NADPH/NADP+ redox couple is differentially regulated by distinct enzymatic machineries at the subcellular level, creating compartmentalized pools that maintain redox homeostasis and support diverse metabolic functions [2] [3]. In the context of increasing interest in metabolic diseases, cancer, and aging, understanding NADPH dynamics has become crucial for therapeutic development [1] [2]. Recent advances in genetically encoded biosensors now enable real-time monitoring of subcellular NADP redox states, revealing unprecedented insights into metabolic regulation [3]. This Application Note examines NADPH's core functions, quantitative contributions of production pathways, and experimental approaches for investigating NADPH dynamics, with emphasis on biosensor methodologies relevant to drug discovery research.
NADPH-Dependent Cellular Functions
Antioxidant Defense Systems
NADPH serves as the essential electron donor for maintaining cellular antioxidant systems, protecting cells against oxidative damage through multiple mechanisms:
- Glutathione Recycling: NADPH is required by glutathione reductase (GSR) to reduce oxidized glutathione (GSSG) to its reduced form (GSH), which then serves as a co-substrate for glutathione peroxidases (GPX) that reduce hydrogen peroxide (H₂O₂) and organic hydroperoxides to water or alcohols [1] [4].
- Thioredoxin System: Thioredoxin reductase (TXNRD) utilizes NADPH to maintain reduced thioredoxin (TRX), which contributes to peroxide scavenging and regulates various redox-sensitive signaling pathways [1].
- Catalase Reactivation: NADPH binds to catalase and reactivates it after oxidative inactivation, enhancing the enzyme's capacity to decompose H₂O₂ [1].
The critical role of NADPH in antioxidant defense is exemplified in oligodendrocyte precursor cells (OPCs), where NADPH depletion induces necrosis through increased superoxide production and glutathione oxidation [5].
Reductive Biosynthesis
NADPH provides reducing equivalents for numerous anabolic pathways essential for cell growth and proliferation:
- Lipid Synthesis: NADPH is required by fatty acid synthase (FASN) for de novo lipogenesis, providing electrons for the reduction reactions during fatty acid elongation [1].
- Cholesterol Synthesis: The mevalonate pathway depends on NADPH as a cofactor for HMG-CoA reductase (HMGCR), the rate-limiting enzyme in cholesterol biosynthesis [1].
- Nucleotide Synthesis: NADPH is needed by dihydrofolate reductase (DHFR) for folate metabolism and the regeneration of tetrahydrofolate, required for de novo purine and thymidylate synthesis [1].
- Amino Acid Metabolism: Iron-sulfur (Fe/S) cluster assembly, essential for various metabolic enzymes including those involved in amino acid biosynthesis, depends on NADPH-derived electrons [1].
Free Radical Generation and Redox Signaling
Paradoxically, NADPH also serves as a substrate for NADPH oxidases (NOX1-5, DUOX1-2), which generate superoxide anions and H₂O₂ as signaling molecules [1]. These NADPH oxidase-derived reactive oxygen species specifically regulate redox-sensitive signaling pathways involved in cellular proliferation, inflammation, and other processes by modulating oncogenes (Src, Ras) and tumor suppressor proteins (TP53, PTEN) [1].
Table 1: Major NADPH-Consuming Pathways and Their Functions
| Pathway/Enzyme | Primary Function | Subcellular Localization |
|---|---|---|
| Glutathione Reductase (GSR) | Reduces GSSG to GSH for antioxidant defense | Cytosol, Mitochondria |
| Thioredoxin Reductase (TXNRD) | Maintains reduced thioredoxin for redox signaling | Cytosol, Mitochondria |
| Fatty Acid Synthase (FASN) | Catalyzes de novo fatty acid biosynthesis | Cytosol |
| HMG-CoA Reductase (HMGCR) | Rate-limiting enzyme in cholesterol synthesis | ER, Cytosol |
| Dihydrofolate Reductase (DHFR) | Regenerates tetrahydrofolate for nucleotide synthesis | Cytosol |
| NADPH Oxidases (NOX) | Generates ROS for signaling and immune function | Plasma Membrane |
Quantitative Analysis of NADPH Production Pathways
The relative contributions of different pathways to NADPH production vary by cell type, metabolic state, and subcellular compartment. Quantitative flux analysis reveals several major NADPH-producing systems:
Pentose Phosphate Pathway (PPP)
The oxidative branch of the PPP is the dominant contributor to cytosolic NADPH in many cell types [6]:
- Glucose-6-Phosphate Dehydrogenase (G6PD): Catalyzes the first committed step of PPP, generating one NADPH molecule per glucose-6-phosphate oxidized [1].
- 6-Phosphogluconate Dehydrogenase (6PGD): Produces a second NADPH molecule while decarboxylating 6-phosphogluconate [1].
Quantitative deuterium tracing studies indicate the PPP contributes 30-50% of total cytosolic NADPH production in proliferating cells, with absolute fluxes of 1-2.5 nmol/μL·h⁻¹ [6]. Inhibition of G6PD sensitizes cells to oxidative stress, demonstrating its critical role in antioxidant defense [5] [6].
Folate-Mediated One-Carbon Metabolism
Surprisingly, serine-driven one-carbon metabolism contributes nearly comparable NADPH to the PPP [6]:
- Methylenetetrahydrofolate Dehydrogenase (MTHFD): Oxidizes methylene-THF to 10-formyl-THF with concomitant reduction of NADP+ to NADPH [6].
- Compartmentalization: Both cytosolic (MTHFD1) and mitochondrial (MTHFD2) isoforms contribute to NADPH homeostasis [6].
Knockdown of MTHFD isozymes decreases cellular NADPH/NADP+ and GSH/GSSG ratios, increasing sensitivity to oxidative stress [6]. This pathway accounts for approximately 40% of NADPH production in some proliferating cells [6].
Other NADPH-Producing Enzymes
- Malic Enzyme (ME1): Oxidatively decarboxylates malate to pyruvate, generating NADPH [1]. Contributes 15-50% of cytosolic NADPH depending on cell type [6].
- Isocitrate Dehydrogenase (IDH1/2): Cytosolic IDH1 and mitochondrial IDH2 catalyze oxidative decarboxylation of isocitrate to α-ketoglutarate, producing NADPH [1] [4].
- NAD+ Kinases (NADK): Phosphorylate NAD+ to NADP+, controlling the substrate pool for NADPH-producing enzymes [1]. Both cytosolic (NADK) and mitochondrial (NADK2) isoforms exist [1].
Table 2: Quantitative Contributions of NADPH Production Pathways in Proliferating Mammalian Cells
| Pathway | Key Enzymes | % Contribution to Cytosolic NADPH | Regulation |
|---|---|---|---|
| Oxidative PPP | G6PD, 6PGD | 30-50% | Substrate availability, NADP+ levels |
| Folate Metabolism | MTHFD1, MTHFD2 | ~40% | Serine availability, purine demand |
| Malic Enzyme | ME1 | 15-50% (cell-type dependent) | Mitochondrial metabolism, glutaminolysis |
| Isocitrate Dehydrogenase | IDH1, IDH2 | Minor contributor (<10%) | Cellular citrate flux, ATP levels |
| Mitochondrial Systems | IDH2, ME3, NNT | Varies by compartment | Mitochondrial redox state |
Diagram 1: Subcellular NADPH Metabolism Network. NADPH is produced through multiple compartmentalized pathways and utilized for antioxidant defense, biosynthesis, and redox signaling.
Experimental Protocols for NADPH Dynamics Analysis
Genetically Encoded Biosensor Applications
The development of genetically encoded biosensors has revolutionized NADPH monitoring, enabling real-time measurement of NADP redox states with subcellular resolution [3]:
NAPstar Biosensor Protocol
Principle: NAPstars are a family of fluorescent protein-based biosensors derived from Peredox-mCherry, with mutated Rex domains that shift specificity from NADH/NAD+ to NADPH/NADP+ [3]. These sensors incorporate circularly permuted T-Sapphire (cpTS) flanked by NADPH-binding domains and an mCherry reference fluorophore.
Experimental Workflow:
Sensor Selection: Choose appropriate NAPstar variant based on dynamic range and affinity:
- NAPstar1 (Kr = 0.004): High affinity for reduced conditions
- NAPstar3 (Kr = 0.026): Intermediate affinity
- NAPstar6 (Kr = 0.41): Lower affinity for oxidized conditions [3]
Subcellular Targeting: Fuse appropriate targeting sequences for compartment-specific expression:
- Cytosolic: No additional sequence
- Mitochondrial: COX VIII presequence
- Nuclear: NLS sequence
- ER: KDEL sequence
Cell Transfection: Transduce cells using preferred method (lentivirus, electroporation, lipofection).
Live-Cell Imaging:
- Maintain cells at 37°C with 5% CO₂ during imaging
- Acquire dual-excitation ratio (400/488 nm) or fluorescence lifetime (FLIM) measurements
- Collect emission at 515 nm for cpTS and 610 nm for mCherry
- Calculate ratio (TS/mCherry) to report NADPH/NADP+ redox state
Calibration: Perform in situ calibration using digitonin permeabilization and NADPH/NADP+ buffers.
Applications: NAPstars have been successfully deployed in yeast, human cell models, and plants, revealing compartment-specific NADPH dynamics during cell division, metabolic shifts, and oxidative stress [3].
iNap Biosensor Protocol
Principle: iNap sensors utilize Rex domains from Bacillus subtilis with cpGFP, responding to NADPH concentration changes [7].
Procedure:
- Express cyto-iNap1 or mito-iNap3 in target cells
- Image using confocal microscopy with 405/488 nm excitation
- Calculate 405/488 ratio to determine NADPH levels
- Normalize using non-responsive iNapc variant [7]
Validation: iNap sensors detected elevated cytosolic NADPH during endothelial cell senescence, demonstrating applications in aging research [7].
Deuterium Tracer Analysis for NADPH Flux Quantification
This metabolic flux approach directly tracks NADPH production pathways using deuterated substrates [6]:
Protocol Steps:
Cell Preparation: Culture cells to 70-80% confluence in appropriate medium.
Isotope Labeling:
- Replace medium with substrate-specific deuterated compounds:
- 1-²H-glucose or 3-²H-glucose for PPP flux
- 2,3,3-²H-serine for folate pathway flux
- 2,3,3,4,4-²H-glutamine for mitochondrial pathways
- Incubate for time course (5 min to 2 hours)
- Replace medium with substrate-specific deuterated compounds:
Metabolite Extraction:
- Rapidly wash cells with ice-cold saline
- Extract with 80% methanol/water at -80°C
- Centrifuge at 16,000 × g for 15 min at 4°C
- Dry supernatant under nitrogen gas
LC-MS Analysis:
- Resuspend samples in LC-MS compatible solvent
- Separate using reverse-phase chromatography (BEH C18 column)
- Analyze with high-resolution mass spectrometer
- Monitor NADP+ (m/z 742.075) and NADPH (m/z 743.079)
Flux Calculation:
- Apply mass isotopomer distribution analysis
- Calculate fractional NADPH labeling: (NADP²H/Total NADPH)
- Apply kinetic isotope effect correction (CKIE ≈ 2.0)
- Use Equation: FractionNADPH from pathway = 2 × (NADP²H/Total NADPH) × (²H-Substrate/Total Substrate)⁻¹ × CKIE [6]
Diagram 2: Complementary Experimental Approaches for NADPH Analysis. Biosensors enable real-time spatial tracking, while deuterium tracing provides quantitative pathway flux measurements.
Research Reagent Solutions
Table 3: Essential Research Tools for NADPH Metabolism Studies
| Reagent Category | Specific Examples | Key Applications | Considerations |
|---|---|---|---|
| Genetically Encoded Biosensors | NAPstar variants (1-7), iNap1, iNap3, SoNar | Real-time NADPH/NADP+ ratio monitoring in live cells | Select based on affinity range (Kr), pH sensitivity, and targeting |
| Deuterated Tracers | 1-²H-glucose, 3-²H-glucose, 2,3,3-²H-serine | Quantitative flux analysis of NADPH production pathways | Account for kinetic isotope effects in calculations |
| Chemical Inhibitors | 6-Aminonicotinamide (6AN), DHEA, Buthionine sulfoximine (BSO) | Pathway inhibition studies; 6AN blocks PPP via G6PD inhibition | 6AN cytotoxicity requires antioxidant rescue controls [5] |
| Enzyme Expression Constructs | MTHFD1/2 overexpression, G6PD shRNA, IDH2 mutants | Genetic manipulation of NADPH production | Mitochondrial targeting requires specific localization sequences |
| Detection Kits & Assays | LC-MS NADPH/NADP+ analysis, LDH cytotoxicity assay, GSH/GSSG assay | Endpoint validation of NADPH status | Rapid quenching essential for accurate redox state preservation |
NADPH serves as the fundamental currency of cellular reducing power, integrating metabolic status with antioxidant defense and biosynthetic capacity. The development of genetically encoded biosensors and sophisticated metabolic tracing approaches has transformed our understanding of NADPH dynamics, revealing remarkable compartmentalization and pathway specialization. The NAPstar biosensor family represents a significant advancement, enabling specific, real-time monitoring of NADP redox states across a 5000-fold range with subcellular resolution [3].
For drug development professionals, targeting NADPH metabolism offers promising therapeutic strategies, particularly in cancer, neurodegenerative diseases, and aging-related conditions. The discovery that folate metabolism contributes substantially to NADPH production reveals new drug targets beyond the classical PPP [6]. Furthermore, the compartment-specific regulation of NADPH pools suggests opportunities for precisely targeted interventions that avoid global metabolic disruption. As research continues to elucidate the complex dynamics of NADPH homeostasis, these insights will undoubtedly inform novel therapeutic approaches for diverse pathological conditions characterized by redox imbalance.
The reduced form of nicotinamide adenine dinucleotide phosphate (NADPH) serves as a fundamental cofactor in metabolic networks, acting as a crucial electron donor for biosynthetic reactions and antioxidant functions in living cells [8] [9]. The NADPH/NADP+ redox couple is differentially regulated by distinct enzymatic machineries at the subcellular level, maintaining a delicate balance essential for cellular function [3]. This redox balance represents a critical interface between metabolism and redox signaling, with implications spanning from cellular oxidative stress responses to the productivity of biotransformation processes in industrial biotechnology [10] [8].
Disruption of this equilibrium creates a cascade of detrimental effects. In vascular diseases, NADPH oxidase-generated reactive oxygen species (ROS) contribute to oxidative stress, endothelial dysfunction, and vascular inflammation that underlie arterial remodelling and atherogenesis [11]. Similarly, in bioproduction, insufficient NADPH regeneration limits the yield of high-value chemicals requiring substantial reducing power, including amino acids, terpenes, and fatty-acid-based fuels [8]. Understanding and monitoring these imbalances through advanced biosensing technologies therefore represents a crucial frontier for both therapeutic intervention and biotechnological advancement.
Pathophysiological Consequences of Redox Imbalance
Oxidative Stress in Metabolic and Vascular Disease
Redox imbalance fundamentally contributes to disease pathophysiology through multiple interconnected mechanisms:
NADH/NAD+ Redox Imbalance in Diabetes: Under diabetic hyperglycemia, NADH becomes overproduced through enhanced flux through glycolytic pathways and the polyol pathway, while NAD+ is depleted through PARP overactivation [10]. The resulting reductive stress initially manifests as NADH accumulation but eventually leads to oxidative stress and damage to macromolecules including DNA, lipids, and proteins [10] [12].
NADPH Oxidase in Vascular Pathology: The NOX1 and NOX2 NADPH oxidases constitute major ROS sources in the artery wall under conditions such as hypertension, hypercholesterolaemia, and diabetes [11]. The generated superoxide anions chemically react with and inactivate nitric oxide (NO), nullifying its vasoprotective functions and promoting formation of the powerful oxidant peroxynitrite [11].
Biomolecular Damage: ROS induce lipid peroxidation in cell membranes, protein carbonylation that compromises enzymatic function, and DNA damage including base modifications and strand breaks, driving mutagenesis and cellular dysfunction [13].
Table 1: Enzymatic Sources and Pathological Consequences of Redox Imbalance
| Enzyme/Pathway | Redox Effect | Pathological Consequences |
|---|---|---|
| Polyol pathway | NADH overproduction, NADPH consumption | Reductive stress, osmotic stress from sorbitol accumulation |
| Mitochondrial complex I | Impaired NAD+ regeneration | Enhanced ROS production, metabolic inhibition |
| NADPH oxidases (NOX1/2) | Superoxide overproduction | Endothelial dysfunction, vascular inflammation |
| PARP overactivation | NAD+ depletion | Cell death, impaired sirtuin function |
Limitations in Bioproduction
In metabolic engineering and bioproduction, NADPH availability frequently constrains process efficiency:
Static Regulation Failures: Traditional static regulation strategies (e.g., promoter engineering, pathway overexpression) often lead to NADPH/NADP+ imbalance because they cannot adjust intracellular NADPH levels in real-time according to varying demands at different culture phases [8].
Cofactor Limitation: The biosynthesis of numerous valuable compounds, including poly-3-hydroxybutyrate (PHB), mevalonate, and fatty acid-based biofuels, is limited by insufficient NADPH regeneration rates and availability [8].
Oxidative Damage to Cells: Under high metabolic loads, insufficient NADPH regeneration compromises the cellular antioxidant defense system, leading to accumulation of ROS and potentially cell death, thereby reducing overall bioprocess yields [8].
Advanced Biosensors for Dynamic Redox Monitoring
Genetically Encoded NADPH/NADP+ Biosensors
Recent advances in genetically encoded biosensors have revolutionized our ability to monitor NADP redox states with subcellular resolution in living cells:
NAPstars Sensor Family: This biosensor family, developed from rational engineering of the Peredox-mCherry scaffold, enables specific, real-time measurements across a broad range of NADPH/NADP+ ratios (approximately 0.001 to 5) [3]. These sensors exhibit high specificity for NADP(H) over NAD(H), with dissociation constants (Kd) for NADPH ranging from 0.9 μM for NAPstar1 to 11.6 μM for NAPstar6 [3].
iNap Sensors: The iNap series represents another class of ratiometric, pH-resistant indicators for NADPH with various affinities (Kd values from ~1.3 μM to ~120 μM) [9]. These permit quantification of separate cytosolic and mitochondrial NADPH pools and reveal NADPH dynamics under oxidative stress depending on glucose availability [9].
NERNST Biosensor: This ratiometric biosensor enables real-time monitoring of NADPH/NADP+ redox status based on a redox-sensitive green fluorescent protein and an NADPH-thioredoxin reductase C module [8].
Table 2: Characteristics of Representative NADPH/NADP+ Biosensors
| Biosensor | Dynamic Range (NADPH/NADP+) | Affinity (Kd) | Key Features | Applications |
|---|---|---|---|---|
| NAPstar1 | 0.001-5 | 0.9 μM (NADPH) | High specificity, compatible with FLIM | Subcellular NADP redox dynamics |
| iNap1 | N/A | ~2.0 μM (NADPH) | pH-resistant, 900% ratio change | Cytosolic NADPH monitoring in mammalian cells |
| iNap3 | N/A | ~25 μM (NADPH) | Moderate affinity | Mitochondrial NADPH quantification |
| NERNST | N/A | N/A | roGFP2-based | NADPH/NADP+ redox status across organisms |
Experimental Protocol: Monitoring NADPH Dynamics in Live Cells Using NAPstar Biosensors
Purpose: To monitor subcellular NADP redox state dynamics in response to oxidative challenge or metabolic perturbations in living cells.
Materials:
- Mammalian cell line (e.g., HeLa, HEK293T) or yeast cells
- NAPstar plasmid DNA (appropriate variant based on desired affinity)
- Transfection reagent (e.g., polyethyleneimine, lipofectamine)
- Confocal fluorescence microscope with temperature and CO₂ control
- Image analysis software (e.g., ImageJ, FIJI)
- Physiological buffer (e.g., HEPES-buffered saline)
Procedure:
- Sensor Expression:
- Transfect cells with NAPstar plasmid using standard transfection protocols.
- Allow 24-48 hours for expression before imaging.
Microscopy Setup:
- Use a confocal microscope with excitation capabilities at ~400 nm and ~500 nm.
- Set emission collection at 515 nm for TS fluorescence and ~610 nm for mCherry.
- Maintain temperature at 37°C with 5% CO₂ for mammalian cells.
Ratiometric Measurement:
- Acquire dual-excitation ratio images (R407/482 for NAPstars).
- Calculate ratio values (TS/mCherry) for each time point.
Experimental Intervention:
- Apply oxidative stress (e.g., H₂O₂ bolus, menadione).
- Induce metabolic perturbations (e.g., glucose deprivation, mitochondrial inhibitors).
- Monitor real-time NADPH dynamics through ratio changes.
Data Analysis:
- Normalize ratio values to baseline (pre-treatment) levels.
- Generate time courses of NADPH/NADP+ ratio changes.
- Compare responses between subcellular compartments.
Troubleshooting:
- For low signal-to-noise ratio, optimize expression level or try higher-affinity sensor variants.
- If pH sensitivity is concerning, utilize the pH-insensitive mCherry normalization.
- For compartment-specific measurements, employ targeted versions (e.g., mitochondrial matrix-targeting sequences).
NADPH Biosensor Experimental Workflow
Dynamic Regulation Strategies for Redox Balance
From Static to Dynamic Metabolic Engineering
Addressing NADPH limitations in bioproduction requires advanced regulation strategies:
Static Regulation Approaches: These include promoter and RBS engineering to direct carbon flux toward NADPH-generating pathways (e.g., pentose phosphate pathway), heterologous expression of NADPH-regenerating enzymes (e.g., isocitrate dehydrogenases), and protein engineering to modify cofactor preference of enzymes [8].
Dynamic Regulation Systems: These sophisticated approaches utilize genetically encoded biosensors to enable real-time monitoring and regulation of intracellular NADP(H) redox status [8]. The transcription factor SoxR biosensor, for instance, specifically responds to NADPH/NADP+ in E. coli, creating a feedback loop for dynamic control [8].
Natural Dynamic Regulation: Some bacteria naturally employ dynamic regulation through metabolic pathway cyclicity. In certain Pseudomonadaceae, the cyclical operation of the Entner-Doudoroff pathway provides varying NADPH supply between growth and stationary phases, optimally supporting different metabolic demands [8].
Experimental Protocol: Implementing Dynamic NADPH Regulation in Microbial Bioproduction
Purpose: To engineer a dynamically regulated system for maintaining NADPH homeostasis during bioproduction of NADPH-intensive compounds.
Materials:
- Microbial chassis (E. coli, yeast)
- NADPH-responsive biosensor (e.g., SoxR-based system, NAPstar)
- Regulatable promoter system (e.g., inducible, CRISPRi)
- Genes for target compound biosynthesis pathway
- Genes for NADPH-regenerating enzymes (e.g., G6PDH, IDH)
- Fermentation equipment
- Analytics (HPLC, MS) for product quantification
Procedure:
- System Construction:
- Integrate NADPH biosensor with regulatable promoter controlling NADPH-regenerating enzymes.
- Incorporate biosynthetic pathway for target compound.
- Verify component functionality in modular fashion.
Characterization:
- Calibrate biosensor response to NADPH fluctuations.
- Determine dynamic range of regulation system.
- Establish correlation between sensor output and product yield.
Bioprocess Optimization:
- Operate bioreactor with controlled feeding strategy.
- Monitor NADPH status in real-time using online or at-line measurements.
- Allow dynamic adjustment of NADPH-regenerating pathway expression.
Performance Evaluation:
- Compare product titers, yields, and productivity between static and dynamic regulation.
- Assess metabolic flux distributions via ¹³C tracing.
- Analyze system robustness over multiple generations.
Troubleshooting:
- If regulation is insufficient, optimize promoter strength or regulatory circuit architecture.
- For metabolic burden, fine-tune expression levels of pathway enzymes.
- If sensor response is slow, consider higher-affinity variants or amplification strategies.
Dynamic NADPH Regulation Circuit
The Scientist's Toolkit: Essential Research Reagents and Solutions
Table 3: Key Research Reagent Solutions for NADPH Redox Studies
| Reagent/Solution | Function | Application Examples |
|---|---|---|
| NAPstar biosensors | Real-time monitoring of NADPH/NADP+ ratio | Live-cell imaging of subcellular NADP redox dynamics |
| iNap sensors | Ratiometric NADPH quantification | Flow cytometry, mitochondrial vs. cytosolic NADPH pools |
| Glucose-6-phosphate dehydrogenase (G6PD) | Rate-limiting enzyme in pentose phosphate pathway | Enhancing NADPH regeneration in bioproduction hosts |
| PARP inhibitors (e.g., 1,5-isoquinolinediol) | Prevent NAD+ depletion under oxidative stress | Studying NAD+ conservation in diabetic models |
| NADK overexpression constructs | Enhance NADP+ synthesis from NAD+ | Manipulating total NADP pool size |
| CD38 inhibitors (e.g., 78c) | Prevent age-related NAD+ decline | Aging research, neurodegenerative disease models |
| SoxR-based regulatory systems | Dynamic control of gene expression | Metabolic engineering for NADPH homeostasis |
Concluding Perspectives
The consequences of NADPH/NADP+ imbalance span from fundamental pathophysiological processes to practical limitations in industrial bioprocesses. The emergence of sophisticated genetically encoded biosensors has transformed our ability to monitor these dynamics with unprecedented spatial and temporal resolution, revealing new insights into cellular redox biology [3] [9]. These tools enable the transition from static to dynamic regulation strategies, offering promising avenues for both therapeutic intervention and biotechnological advancement.
Future directions will likely focus on refining these dynamic control systems, enhancing biosensor performance characteristics, and integrating multi-omic approaches to comprehensively understand redox regulation networks. The continued development of this toolkit will be essential for addressing the dual challenges of oxidative stress in disease and metabolic limitations in bioproduction, ultimately advancing both human health and sustainable manufacturing paradigms.
The regulation of intracellular redox states, particularly the balance between NADPH and NADP+, is fundamental to cellular metabolism, antioxidant defense, and reductive biosynthesis. Traditional approaches to studying and engineering these systems have relied heavily on static regulation strategies and endpoint biochemical assays. However, these methods possess inherent limitations that obscure the dynamic nature of metabolic processes. This application note delineates the critical shortcomings of traditional methodologies and champions the integration of genetically encoded biosensors for real-time, subcellular monitoring of NADP(H) dynamics. We present experimental protocols and key reagents that empower researchers to overcome these limitations, thereby enabling precise dynamic regulation of metabolic pathways for advanced bioproduction and therapeutic development.
The NADPH/NADP+ redox couple constitutes a central metabolic node, providing essential reducing power for biosynthetic reactions, cellular antioxidant defense, and redox signaling [8]. The NADPH/NADP+ ratio is differentially regulated across subcellular compartments—cytosol, mitochondria, and other organelles—each maintaining distinct pool sizes and redox states [14]. Precise maintenance of this balance is critical; deviation can lead to oxidative stress, metabolic dysfunction, and reduced productivity in engineered systems.
Conventional metabolic engineering has primarily employed static regulation strategies, such as constitutive overexpression or knockout of genes, to modulate NADPH regeneration and consumption [8]. The efficacy of these strategies is assessed using traditional biochemical assays. However, a growing body of evidence indicates that these conventional approaches are insufficient for capturing and controlling the dynamic, compartmentalized nature of central redox metabolism, necessitating a paradigm shift toward real-time monitoring and dynamic regulation.
Limitations of Traditional Methods
Shortcomings of Static Regulation Strategies
Static regulation strategies, while straightforward to implement, fail to respond to the changing metabolic demands of a cell over time, often leading to metabolic imbalance.
- Metabolic Imbalance and Cytotoxicity: Constitutive overexpression of pathways to enhance NADPH supply (e.g., the pentose phosphate pathway) often disrupts the NADPH/NADP+ equilibrium. This imbalance can impair cell growth and viability, ultimately limiting the yield of target compounds, such as fatty acids, terpenes, and amino acids [8]. In bioproduction, the accumulation of cytotoxic intermediates like cadaverine can arrest growth when using static control [15].
- Lack of Temporal Control: Static strategies cannot adjust to the varying NADPH requirements at different growth phases (e.g., growth phase vs. production phase). This inflexibility results in suboptimal resource allocation and fails to protect cells from transient metabolic stresses [8].
- Inability to Maintain Redox Homeostasis: The cellular redox state is a dynamic property. Static interventions are inherently incapable of fine-tuning the NADPH/NADP+ ratio in real-time to maintain homeostasis under fluctuating environmental conditions, such as oxidative stress or changes in nutrient availability [8].
Pitfalls of Endpoint Biochemical Assays
Traditional methods for quantifying NADP(H) levels provide only a snapshot in time and lack the spatial resolution to understand subcellular metabolic heterogeneity.
- Destructive and Low-Temporal Resolution: Techniques like chromatography and mass spectrometry require cellular lysis, destroying the spatial information and making it impossible to track metabolic kinetics in the same cell population over time [14].
- Inability to Distinguish NADPH from NADH: NAD(P)H autofluorescence measurement is a non-invasive method but suffers from low sensitivity, potential cell damage from ultraviolet irradiation, and a critical inability to distinguish between the fluorescence signals of NADH and NADPH due to their nearly identical spectra [14].
- Low Spatial Resolution and Technical Complexity: While Fluorescence Lifetime Imaging (FLIM) can potentially differentiate bound NADH from NADPH, it is a technically demanding methodology not readily accessible to most laboratories. Other techniques, like in situ magnetic resonance imaging for NAD+, are prohibitively complex for routine use [14].
- Artifact-Prone Semisynthetic Probes: Semisynthetic FRET probes (e.g., NAD-Snifit) require extensive washing to remove excess dye, a process that can introduce artifacts and is poorly suited for dynamic developmental or long-term studies [14].
Table 1: Quantitative Comparison of NADP(H) Analysis Methods
| Method | Temporal Resolution | Spatial Resolution | Destructive? | Can Distinguish NADPH & NADH? | Key Limitation |
|---|---|---|---|---|---|
| LC-MS/Chromatography | Endpoint (Hours) | None (Population Average) | Yes | Yes | No live-cell monitoring [14] |
| NAD(P)H Autofluorescence | Seconds-Minutes | Subcellular | No | No | Low sensitivity, non-specific [14] |
| FLIM | Seconds | Subcellular | No | Partially (for bound states) | Technically complex [14] |
| Semisynthetic Probes (e.g., NADP-Snifit) | Minutes | Subcellular | No | Yes | Requires washing, prone to artifacts [14] |
| Genetically Encoded Biosensors (e.g., NAPstars) | Seconds | Subcellular | No | Yes | Requires genetic engineering [3] |
Biosensors as a Solution for Real-Time Monitoring
Genetically encoded biosensors represent a revolutionary technology that overcomes the limitations of traditional methods, enabling specific, sensitive, and non-invasive monitoring of metabolite dynamics in living cells and in vivo.
These biosensors typically consist of a sensing domain derived from a bacterial transcriptional repressor (e.g., Rex) that binds specifically to NADP(H), fused to one or more fluorescent proteins (e.g., cpT-Sapphire, mCherry). Upon ligand binding, a conformational change in the sensing domain alters the fluorescence properties of the reporter, providing a quantifiable ratiometric readout of the NADPH/NADP+ ratio [3].
The NAPstar Family of NADP(H) Biosensors
The recently developed NAPstar family of biosensors exemplifies the power of this technology. Derived from the NAD+ sensor Peredox, NAPstars incorporate specific mutations to shift binding preference towards NADP(H) [3].
Table 2: Characteristics of Select NAPstar Biosensor Variants
| Biosensor Variant | Apparent Kr (NADPH/NADP+) | Dynamic Range | Key Application Note |
|---|---|---|---|
| NAPstar1 | ~0.006 | ~2.5 | Highest affinity; ideal for detecting low ratio values [3] |
| NAPstar3 | ~0.03 | ~2.5 | Balanced affinity; used in yeast, plants, mammalian cells [3] |
| NAPstar6 | ~0.08 | ~2.5 | Lower affinity; suitable for compartments with highly reduced NADP pools [3] |
| NAPstarC | N/A (Control) | N/A | Non-binding control for validating signal specificity [3] |
Key advantages of the NAPstar biosensors include:
- High Specificity: Significant affinity for NADP(H) over NAD(H), with Kd(NADH) values one to two orders of magnitude higher than Kd(NADPH) [3].
- Ratiometric Output: The fusion of a sensing fluorescent protein (cpT-Sapphire) to a reference fluorescent protein (mCherry) allows for ratio-metric measurement, minimizing artifacts from changes in sensor concentration or expression level.
- Compartment-Specific Targeting: They can be genetically targeted to specific subcellular locations (e.g., cytosol, mitochondria) to reveal compartmentalized redox dynamics [3].
- Bona Fide Redox State Reporting: NAPstars are predominantly sensitive to the NADPH/NADP+ ratio rather than the absolute concentration of either molecule, making them ideal reporters of redox state [3].
Experimental Protocols
Protocol: Monitoring NADP(H) Redox Dynamics in Cultured Mammalian Cells Using NAPstars
Purpose: To monitor real-time changes in the cytosolic NADPH/NADP+ ratio in response to an oxidative challenge.
Reagents and Equipment:
- Cell Line: HEK-293T or other relevant mammalian cell line.
- Plasmid: pDNA-NAPstar3 (cytosolic), available from Addgene (#XXXXX).
- Transfection Reagent: Polyethylenimine (PEI) or lipofectamine.
- Imaging Buffer: Hanks' Balanced Salt Solution (HBSS) or phenol-red free culture medium.
- Stimuli: Dihydroethidium (DHE) or Menadione as a source of oxidative stress.
- Microscope: Confocal or widefield fluorescence microscope equipped with:
- Excitation Lasers/Lights: 405 nm (for cpT-Sapphire), 560 nm (for mCherry).
- Emission Filters: 515/30 nm (for cpT-Sapphire), 630/50 nm (for mCherry).
Procedure:
- Cell Seeding and Transfection:
- Seed cells onto poly-D-lysine-coated glass-bottom imaging dishes at 50-70% confluency.
- 24 hours post-seeding, transfert cells with the pDNA-NAPstar3 construct using your preferred transfection method.
- Incubate for 24-48 hours to allow for sufficient biosensor expression.
Microscope Setup and Calibration:
- Pre-warm the microscope stage to 37°C with 5% CO₂ supplementation.
- Set up a time-lapse experiment with sequential imaging of both channels to avoid bleed-through.
- For cpT-Sapphire: Excite at 405 nm, collect emission at 515/30 nm.
- For mCherry: Excite at 560 nm, collect emission at 630/50 nm.
Baseline Acquisition:
- Replace the culture medium with pre-warmed imaging buffer.
- Acquire images every 30-60 seconds for a 5-minute baseline period to establish the initial ratio (R₀).
Oxidative Challenge:
- Without moving the field of view, carefully add the oxidative stressor (e.g., 100 µM DHE) directly to the imaging buffer. Mix gently.
- Continue time-lapse imaging for an additional 20-40 minutes.
Data Analysis:
- For each time point, calculate the background-subtracted fluorescence intensity for both cpT-Sapphire (F₁) and mCherry (F₂) channels in your region of interest (single cell or cytoplasm).
- Compute the ratio R = F₁ / F₂ for each time point.
- Normalize the ratios to the average baseline ratio (R₀) and plot as R/R₀ over time. A decrease in the ratio indicates a shift towards a more oxidized NADP pool.
Protocol: Dynamic Regulation of a Biosynthetic Pathway Using a Transcription Factor-Based Biosensor
Purpose: To dynamically upregulate a NADPH-consuming biosynthetic pathway in response to high intracellular NADPH levels, thereby maintaining redox balance and increasing product yield.
Reagents and Equipment:
- Strain: Engineered E. coli production strain.
- Biosensor Circuit: Plasmid containing the SoxR-based biosensor, which activates expression in response to a more reduced NADP pool [8].
- Fermentation Equipment: Shake flasks or bioreactors.
Procedure:
- Strain Engineering:
- Identify a key gene in your target biosynthetic pathway that is limiting for flux (e.g., a reductase).
- Clone this gene under the control of the SoxR-responsive promoter (PsoxR), creating an output module.
- Co-transform or integrate the biosensor (SoxR) and output modules into your production host.
Fermentation and Monitoring:
- Inoculate the engineered strain into a suitable production medium in a bioreactor.
- Maintain optimal growth conditions (pH, temperature, dissolved oxygen).
- Periodically sample the culture to measure both cell density (OD₆₀₀) and product titer (e.g., via HPLC).
Validation of Dynamic Control:
- Compare the final product titer and cell growth of the dynamically regulated strain against two control strains:
- Control 1: A strain with the output gene constitutively expressed.
- Control 2: A strain with the output gene deleted or not expressed.
- The dynamically regulated strain should demonstrate superior performance, with a higher product titer and improved growth compared to the constitutive control, as seen in the cadaverine biosynthesis example [15].
- Compare the final product titer and cell growth of the dynamically regulated strain against two control strains:
The Scientist's Toolkit: Key Research Reagent Solutions
Table 3: Essential Reagents for NADP(H) Biosensor Research
| Item | Function | Example & Specification |
|---|---|---|
| Genetically Encoded Biosensor Plasmids | Core reagent for live-cell imaging of NADP(H) | NAPstar series (e.g., NAPstar3 for cytosol, NAPstar3-mito for mitochondria); Available from Addgene [3]. |
| Oxidative Stressors | To perturb and test the NADP(H) redox system | Dihydroethidium (DHE), Menadione, Hydrogen Peroxide (H₂O₂); prepare fresh stock solutions in DMSO or water [3]. |
| Fluorescent Protein Antibodies | To verify biosensor expression and localization | Anti-GFP antibody (may cross-react with cpT-Sapphire), Anti-RFP antibody (for mCherry); validate for use in your host species. |
| Mammalian Expression Vectors | For biosensor delivery into mammalian cells | pcDNA3.1, pLVX; ensure the vector uses a strong, constitutive promoter (e.g., CMV). |
| Microscopy Calibration Slides | To align and calibrate microscope channels for accurate ratiometry | Multi-channel fluorescent calibration slides, 0.17µm thickness. |
Visualizing Workflows and Signaling Pathways
NADP(H) Metabolism and Biosensor Mechanism
NADPH Metabolism and Biosensor Mechanism
Static vs. Dynamic Regulation Workflow
Static vs Dynamic Regulation Workflow
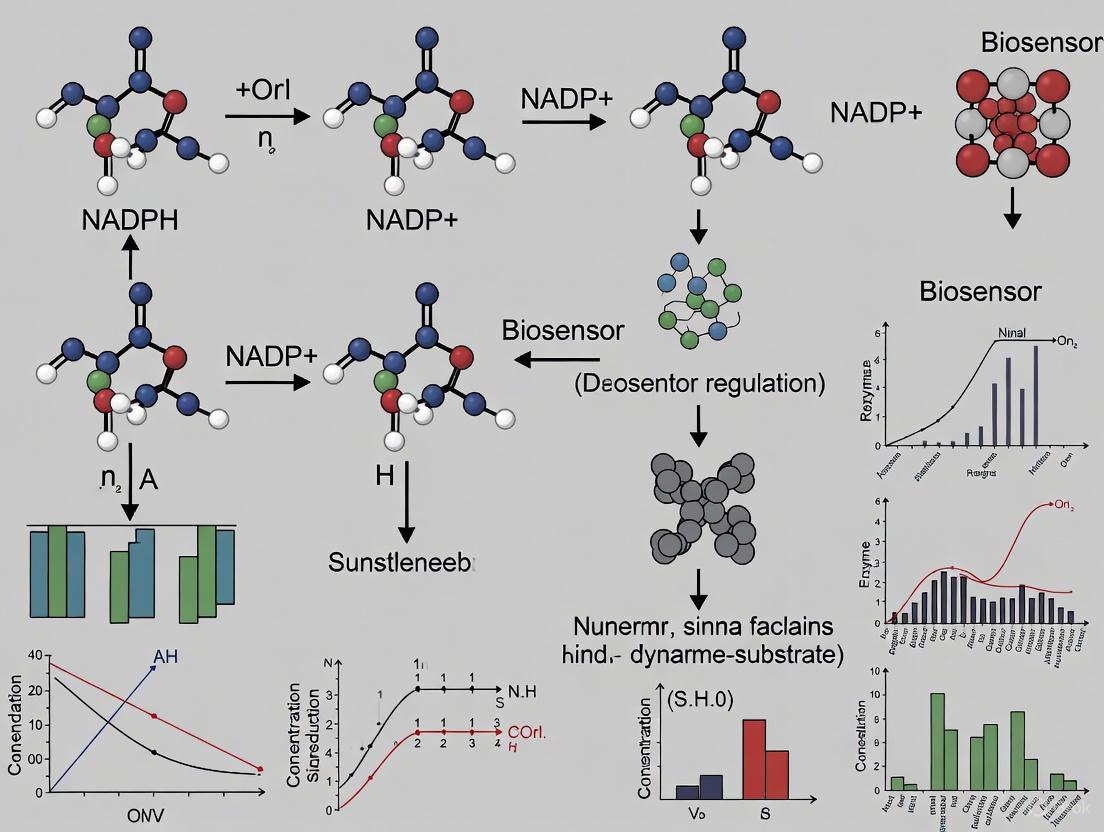
Cellular metabolism is a dynamic network of biochemical reactions essential for life. Understanding the spatial and temporal regulation of metabolites like the NADPH/NADP+ redox couple is crucial, as they constitute central metabolic redox couples involved in numerous cellular functions, from biosynthetic reactions to antioxidant defense [3] [14]. Traditional biochemical methods, such as chromatography and mass spectrometry, require cellular lysis, providing only static snapshots and disrupting the native cellular context [14]. The autofluorescence of NADPH itself is limited by low sensitivity, the inability to distinguish it from NADH, and cell injury caused by ultraviolet irradiation [14].
Genetically encoded fluorescent biosensors have emerged as a revolutionary technology, overcoming these limitations by enabling non-invasive, real-time monitoring of metabolites in living cells and in vivo with high spatiotemporal resolution [16] [14] [17]. These biosensors are engineered proteins that change their fluorescent properties upon binding a target analyte or in response to enzymatic activity. Their genetic encodability allows for precise targeting to specific cell types or subcellular compartments, facilitating the study of metabolic processes with unprecedented detail [18]. This article introduces the core principles of these biosensors and provides detailed protocols for their application, particularly focusing on the dynamic regulation of the NADPH/NADP+ balance.
The Biosensor Toolkit: Design Principles and Key Players
Fundamental Sensing Strategies
Genetically encoded biosensors primarily utilize two reporting mechanisms, each with distinct advantages [19] [17]:
- Förster Resonance Energy Transfer (FRET)-based Biosensors: These consist of two fluorescent proteins (a donor and an acceptor) linked by a sensing domain that undergoes a conformational change upon analyte binding. This change alters the distance or orientation between the fluorophores, modulating FRET efficiency, which is typically measured as a ratio of acceptor-to-donor emission [19]. An example is the NADP+ biosensor, NADPsor, which uses CFP and YFP [20].
- Intensiometric Biosensors: These single-fluorophore sensors incorporate a circularly permuted fluorescent protein (cpFP) into the sensing domain. Analyte-induced conformational changes directly affect the fluorophore's chromophore environment, leading to a change in fluorescence intensity [21] [17]. The GCaMP calcium sensors and the recently developed red lactate sensor R-eLACCO2.1 are prominent examples [21].
Other strategies include monitoring biosensor translocation within the cell or utilizing fluorescence lifetime imaging microscopy (FLIM), which is independent of concentration and excitation intensity, as demonstrated with R-eLACCO2.1 [21] [19].
A Focus on NADPH/NADP+ Biosensors
The NADPH/NADP+ redox couple is differentially regulated across subcellular compartments and is central to redox homeostasis and anabolic metabolism [3] [14]. The following table summarizes key genetically encoded biosensors for tracking this balance.
Table 1: Genetically Encoded Biosensors for NADPH and NADP+ Redox State
| Biosensor Name | Target | Reporting Mechanism | Key Characteristics | Dynamic Range / Affinity |
|---|---|---|---|---|
| NAPstars [3] | NADPH/NADP+ Ratio | Ratiometric (cpT-Sapphire / mCherry) | Real-time, specific measurements across a broad range of NADP redox states; compatible with FLIM. | Kr (NADPH/NADP+) from ~0.001 to 5 |
| NADPsor [20] | NADP+ | FRET (CFP/YFP) | Highly specific to NADP+; optimized via computational protein redesign. | Detection limit of 1 μM; broad range to 10 mM |
| iNaps [3] | NADPH | Intensiometric (single cpFP) | Requires dimerization for function; sensitive to sensor concentration. | Kd(NADPH) in the low micromolar range |
| SoNar [14] | NAD+/NADH & NADP+/NADPH | Intensiometric (cpFP) | High sensitivity but can cross-react with both NADH and NADPH. | Sensitive to cellular redox state |
| Peredox [14] | NAD+/NADH Ratio | Ratiometric (cpT-Sapphire / mCherry) | Chassis for developing NAPstars; primarily measures NAD redox state. | Kd(NADH) = 1.2 μM |
The NAPstar family represents a significant recent advancement. Derived from the NAD+ sensor Peredox, NAPstars are engineered with mutations that switch the specificity of the bacterial Rex domain from NADH to NADPH, creating sensors that report the bona fide NADPH/NADP+ ratio rather than just the concentration of one species [3]. Their design, featuring two Rex domains within a single polypeptide, minimizes concentration-dependent effects and provides a stable, ratiometric readout.
Diagram: Logical framework for selecting and applying NADP(H) biosensors in a research workflow.
Experimental Protocols: From Setup to Imaging
This section provides a detailed methodology for employing NADPH/NADP+ biosensors, using the NAPstar family as a prime example.
Protocol: Monitoring Cytosolic NADP Redox State in Mammalian Cells
Objective: To express NAPstar biosensors in mammalian cells and measure dynamic changes in the NADPH/NADP+ ratio in response to oxidative challenge.
Research Reagent Solutions & Essential Materials
Table 2: Key Reagents and Materials for Biosensor Imaging
| Item | Function / Explanation | Example / Note |
|---|---|---|
| NAPstar Plasmid DNA [3] | Genetically encodes the biosensor. | Select variant (e.g., NAPstar1 for high affinity). Obtain from academic depository (Addgene). |
| Cell Culture Vessels | Live-cell imaging. | Glass-bottom dishes are essential for high-resolution microscopy. |
| Transfection Reagent | Introduces plasmid DNA into cells. | Use reagents compatible with your cell line (e.g., lipofectamine, PEI). |
| Fluorescence Microscope | Detects biosensor signal. | Requires capabilities for ratiometric imaging (e.g., CFP/YFP filters or hyperspectral detection). |
| Excitation Light Sources | Activates the fluorophores. | LEDs or lasers at 400-410 nm (T-Sapphire) and 560-580 nm (mCherry). |
| Emission Filters | Isolates biosensor emission. | Bandpass filters at 515-535 nm (T-Sapphire) and 610-650 nm (mCherry). |
| Pharmacological Agents | Modulate cellular redox state. | Hydrogen peroxide (H₂O₂) to induce oxidative stress; DPI to inhibit NADPH oxidases. |
Procedure:
Sensor Selection and Cloning:
- Select the appropriate NAPstar variant based on your expected NADPH/NADP+ ratio range. NAPstar1 (Kr ~0.01) is suitable for more reduced conditions, while NAPstar6 (Kr ~1) is better for more oxidized pools [3].
- Subclone the NAPstar cDNA into a mammalian expression vector with a strong, ubiquitous promoter (e.g., CMV, CAG).
Cell Culture and Transfection:
- Culture your chosen mammalian cells (e.g., HEK293, HeLa, or primary neurons) according to standard protocols.
- At 50-70% confluency, transfect cells with the NAPstar plasmid using a suitable transfection reagent. Include untransfected controls to assess autofluorescence.
- Allow 24-48 hours for sufficient biosensor expression and maturation before imaging.
Microscope Setup and Calibration:
- Use an inverted fluorescence microscope equipped with stable light sources and sensitive cameras (e.g., EM-CCD or sCMOS).
- Configure the system for sequential dual-excitation ratiometric imaging:
- Excitation 1: 405 nm for T-Sapphire (TS).
- Emission 1: 535/30 nm bandpass filter.
- Excitation 2: 560 nm for mCherry (mC).
- Emission 2: 620/60 nm bandpass filter.
- Minimize exposure time and light intensity to prevent phototoxicity and photobleaching.
Image Acquisition and Baseline Measurement:
- Transfer the culture dish to the microscope stage maintained at 37°C and 5% CO₂.
- Identify healthy, transfected cells exhibiting bright fluorescence in both channels.
- Acquire time-lapse images of both fluorescence channels at a low frequency (e.g., every 30 seconds) to establish a stable baseline TS/mC ratio.
Stimulation and Dynamic Recording:
- Without moving the field of view, carefully add the oxidizing agent (e.g., 100 µM - 1 mM H₂O₂) to the culture medium.
- Continue time-lapse acquisition, increasing the frequency if capturing rapid transients. The TS/mC ratio will decrease as the NADP pool becomes more oxidized (less NADPH bound) [3].
Data Analysis:
- For each time point, calculate the background-subtracted fluorescence intensity ratio (R = TS / mC).
- Normalize the ratios to the initial baseline average (R/R₀) to visualize relative changes.
- The mCherry signal serves as an internal reference for biosensor concentration and cell thickness, correcting for non-specific intensity fluctuations.
Protocol: Validating NADP Redox State with the Glutathione System
Objective: To apply NAPstars in combination with selective impairment of antioxidative pathways to delineate electron flux.
Procedure:
- Follow steps 1-4 from the previous protocol to establish cells expressing NAPstar and acquire a baseline ratio.
- Inhibit the Glutathione System: Treat cells with buthionine sulfoximine (BSO, 100 µM for 18-24 hours) to deplete cellular glutathione by inhibiting γ-glutamylcysteine synthetase.
- Induce Oxidative Stress: Apply a bolus of H₂O₂ (e.g., 200 µM) to the BSO-pre-treated cells and control cells.
- Image and Compare: Acquire time-lapse images as described. Research using this approach has revealed that in BSO-treated cells, the NADP pool remains more oxidized for a longer duration after H₂O₂ challenge, uncovering the glutathione system as the primary mediator of antioxidative electron flux in response to acute oxidative challenge across diverse eukaryotes [3]. This demonstrates the power of biosensors in dissecting specific metabolic pathways.
Diagram: The experimental workflow for biosensor-based metabolic imaging.
Advanced Applications and Visualization in Metabolic Research
The high spatial and temporal resolution of genetically encoded biosensors enables the investigation of complex metabolic phenomena. Key applications illuminated by these tools include:
- Uncovering Metabolic Compartmentalization: By targeting biosensors to specific organelles (e.g., mitochondria, cytosol), researchers can observe independent NADPH dynamics in different subcellular compartments, revealing a previously unappreciated level of metabolic regulation [3] [18].
- Revealing Metabolic Oscillations and Dynamics: NAPstars have been used to uncover cell cycle-linked oscillations in the NADP redox state in yeast and illumination-dependent dynamics in plant leaves, demonstrating that metabolism is highly dynamic and intertwined with cellular programs [3].
- Multiplexing with Other Signaling Pathways: The development of spectrally orthogonal biosensors, such as the red fluorescent lactate sensor R-eLACCO2.1, allows for dual-color imaging with green biosensors like GCaMP (Ca²⁺). This enables the simultaneous observation of extracellular lactate dynamics and neural activity in the somatosensory cortex of awake, behaving mice, providing direct insight into neuro-metabolic coupling [21].
- Dissecting Antioxidant Pathways: As outlined in the validation protocol, biosensors are instrumental in delineating the contributions of parallel systems, such as the glutathione and thioredoxin pathways, in maintaining redox homeostasis during oxidative stress [3].
Genetically encoded biosensors like the NAPstar family have fundamentally transformed our ability to monitor central metabolic pathways, such as the NADPH/NADP+ balance, in their native, living context. They provide quantitative, real-time data with subcellular resolution, moving beyond the static snapshots offered by traditional destructive methods.
Future developments in this field will focus on expanding the color palette of biosensors for enhanced multiplexing, improving their brightness and dynamic range, and engineering new sensing domains to target a wider array of metabolites. The integration of biosensors with advanced microscopy techniques, including super-resolution and in vivo deep-tissue imaging, will continue to illuminate the intricate spatiotemporal dynamics of metabolism, driving forward discoveries in fundamental biology and the development of novel therapeutics for metabolic diseases.
A Practical Guide to NADP(H) Biosensors: From Design to Real-World Applications
The real-time monitoring of intracellular metabolite dynamics is a cornerstone of modern metabolic engineering. For researchers focused on the critical redox balance between NADPH and NADP+, genetically encoded biosensors provide indispensable tools for live-cell imaging and high-throughput screening. These biosensors transduce biochemical signals into quantifiable optical outputs, enabling the dynamic tracking of metabolic fluxes without disrupting cellular integrity. Among the diverse architectures, FRET-based, single fluorescent protein (FP)-based, and transcription factor-based biosensors have emerged as the most prominent designs, each offering distinct advantages for specific experimental applications. This article details the operational principles, standard protocols, and key reagents for these three major biosensor classes, providing a framework for their application in NADPH/NADP+ homeostasis research.
FRET-Based Biosensors
Principle of Operation
Förster Resonance Energy Transfer (FRET)-based biosensors function as molecular rulers, measuring distance changes through non-radiative energy transfer between two fluorophores. The core mechanism involves a donor fluorophore in its excited state transferring energy to an adjacent acceptor fluorophore through dipole-dipole interactions when their separation is within 1-10 nm [22] [23]. This energy transfer results in a decrease in donor emission and an increase in acceptor emission, which can be precisely quantified. The efficiency of FRET (EFRET) exhibits an inverse sixth-power relationship with the distance (R) between the donor and acceptor, as described by the equation EFRET = R06/(R06 + R6), where R0 is the Förster distance at which 50% energy transfer occurs [22]. This exquisite distance sensitivity allows FRET biosensors to detect subtle conformational changes in sensing domains induced by analyte binding, making them ideal for monitoring real-time fluctuations in cofactor levels such as NADPH/NADP+.
Key Design Parameters
Table 1: Critical Parameters for FRET Biosensor Design
| Parameter | Description | Optimal Range/Value | Impact on Performance |
|---|---|---|---|
| Distance (R) | Separation between donor and acceptor fluorophores | 1-10 nm [22] | Inverse sixth-power relationship with FRET efficiency |
| Spectral Overlap (J(λ)) | Integral of donor emission and acceptor absorption spectra | High overlap required [22] | Determines R0 value; greater overlap enables longer-range FRET |
| Orientation Factor (κ²) | Relative dipole orientation between donor and acceptor | 0-4 (theoretical); ~2/3 for isotropic rotation [24] | Poor alignment can eliminate FRET signal even at optimal distances |
| Förster Distance (R0) | Distance for 50% FRET efficiency | 3-6 nm for most FP pairs [24] | Characteristic for each donor-acceptor pair; determines operational range |
Experimental Protocol: FRET Biosensor Imaging for NADPH/NADP+ Dynamics
Purpose: To monitor real-time NADPH/NADP+ ratios in living cells using a FRET-based biosensor.
Materials:
- Genetically encoded FRET biosensor (e.g., specific for NADPH/NADP+ redox state)
- Appropriate cell line (e.g., HEK293, HeLa, or relevant microbial systems)
- Culture medium and supplements
- Ligands/effectors for perturbation studies
- Confocal microscope or fluorescence plate reader with dual-emission capability
- Image analysis software (e.g., ImageJ, MetaMorph)
Procedure:
Biosensor Expression:
- Transfect cells with the FRET biosensor construct using standard methods (lipofection, electroporation, viral transduction).
- Allow 24-48 hours for expression, ensuring proper subcellular targeting if required.
Image Acquisition:
- Maintain cells at 37°C and 5% CO2 during imaging.
- Excite the donor fluorophore at its optimal wavelength (e.g., 433 nm for CFP).
- Collect emissions simultaneously or sequentially for both donor (e.g., 475 nm for CFP) and acceptor (e.g., 527 nm for YFP) channels.
- Acquire time-series images at appropriate intervals (seconds to minutes) depending on the biological process.
FRET Quantification:
- Calculate FRET ratio as (acceptor emission intensity)/(donor emission intensity).
- Correct for background fluorescence, bleed-through, and direct acceptor excitation.
- For more precise measurements, utilize fluorescence lifetime imaging (FLIM) to detect donor lifetime changes induced by FRET [22].
Data Analysis:
- Normalize FRET ratios to baseline values.
- Correlate ratio changes with experimental perturbations.
- Convert ratio values to NADPH/NADP+ concentrations using appropriate calibration curves.
Figure 1: FRET Biosensor Working Principle. Analyte binding induces a conformational change that alters the distance between donor and acceptor fluorophores, modulating FRET efficiency quantified as an emission ratio.
Single Fluorescent Protein Biosensors
Principle of Operation
Single FP biosensors utilize circularly permutated fluorescent proteins (cpFPs) where the original N- and C-termini are linked with a short peptide and new termini are created at another location in the β-barrel structure [17]. This rearrangement positions the chromophore sensitive to conformational changes in fused sensing domains. When analytes such as NADPH or NADP+ bind to the sensing domain, structural rearrangements alter the chromophore's protonation state or the surrounding electrostatic environment, resulting in measurable changes in fluorescence intensity or excitation spectrum [17] [25]. Single FP biosensors offer advantages including simpler implementation, easier targeting to subcellular compartments, and compatibility with multi-color imaging when combined with other biosensors.
Experimental Protocol: Ratiometric Imaging with Single FP Biosensors
Purpose: To detect NADPH/NADP+ ratios using a single FP biosensor with ratiometric capabilities.
Materials:
- Single FP biosensor (e.g., cpFP-based NADPH/NADP+ sensor)
- Appropriate cell line
- Culture medium
- Confocal microscope or ratiometric-capable imaging system
- Image analysis software
Procedure:
Biosensor Expression and Validation:
- Transfect cells with the single FP biosensor construct.
- Verify proper expression and localization via fluorescence microscopy.
Dual-Excitation Imaging:
- For intensity-based sensors: Image at single excitation/emission wavelengths.
- For ratiometric sensors: Acquire images using two alternating excitation wavelengths (e.g., 400 nm and 490 nm for a single emission at 535 nm).
- Maintain consistent acquisition parameters throughout the experiment.
Data Processing:
- For ratiometric sensors, calculate the emission ratio (excitation₁/excitation₂).
- Normalize ratios to baseline values.
- Convert ratio values to analyte concentrations using in situ calibration curves generated with known concentrations of NADPH and NADP+.
Specific Considerations:
- Account for potential pH sensitivity, as some FP chromophores are pH-sensitive.
- Control for biosensor expression levels, as intensity-based measurements are concentration-dependent.
- Perform photostability tests to ensure minimal bleaching during time-lapse experiments.
Figure 2: Single FP Biosensor Mechanism. Circular permutation enables fusion of sensing domains that modulate the chromophore environment upon analyte binding, altering fluorescence properties.
Transcription Factor-Based Biosensors
Principle of Operation
Transcription factor-based biosensors (TFBs) utilize allosteric transcription factors (aTFs) that undergo conformational changes upon binding specific ligands, thereby regulating transcription of reporter genes [26] [27]. In their native state, aTFs bind operator sequences to repress or activate transcription of downstream genes. Ligand binding induces conformational changes that alter DNA binding affinity, resulting in increased or decreased expression of reporter proteins such as fluorescent proteins or enzymes [26]. TFBs are particularly valuable for high-throughput screening applications in metabolic engineering, enabling selection of microbial strains with desired NADPH/NADP+ balance or enzymatic activities without requiring specialized equipment.
Advanced Engineering Approaches
Recent advances in TFB engineering have dramatically expanded their utility. The Sensor-seq platform enables high-throughput design of aTFs for non-native ligands through phylogeny-guided sequence diversification and RNA barcoding coupled to deep sequencing [27]. This approach allows screening of thousands of aTF variants against target ligands, identifying rare functional biosensors with desired specificity and dynamic range. For NADPH/NADP+ sensing, such platforms facilitate the development of customized biosensors even when natural aTFs for these cofactors are unavailable or suboptimal.
Experimental Protocol: TFB-Mediated High-Throughput Screening
Purpose: To screen microbial libraries for NADPH/NADP+ balance using transcription factor-based biosensors.
Materials:
- Microbial strain library (e.g., E. coli, yeast)
- TFB specific for NADPH/NADP+ or related metabolites
- Culture medium and inducers if required
- Flow cytometer or fluorescence-activated cell sorter (FACS)
- Microplate readers for bulk measurements
- Lysis buffers for enzymatic assays if using non-fluorescent reporters
Procedure:
Biosensor Integration:
- Stably integrate the TFB construct into the host genome or maintain on plasmids.
- Ensure the reporter gene (e.g., GFP, RFP) is under control of the aTF-responsive promoter.
Library Cultivation and Induction:
- Grow microbial library under selective conditions.
- Apply metabolic perturbations or chemical treatments to modulate NADPH/NADP+ ratios.
- Allow sufficient time for reporter protein accumulation (several hours to overnight).
Screening and Selection:
- For fluorescent reporters: Analyze cells via flow cytometry or FACS.
- Gate populations based on fluorescence intensity corresponding to desired NADPH/NADP+ levels.
- Sort high- and low-fluorescence populations for further analysis or validation.
- For enzymatic reporters: Perform bulk lysis and spectrophotometric/fluorometric assays.
Validation and Iteration:
- Validate sorted clones for desired metabolic phenotypes.
- Re-screen enriched populations if necessary.
- Sequence genomic regions of interest to identify mutations responsible for metabolic changes.
Figure 3: TFB Operational Mechanism. Ligand binding induces conformational changes in the allosteric transcription factor, altering its DNA binding affinity and modulating reporter gene expression.
Research Reagent Solutions
Table 2: Essential Research Reagents for Biosensor Development and Application
| Reagent Category | Specific Examples | Function/Application | Key Characteristics |
|---|---|---|---|
| Fluorescent Proteins | CFP, YFP, GFP, RFP, cpFPs [17] | FRET pairs, single FP biosensor scaffolds | Spectral properties, brightness, photostability, pH sensitivity |
| Sensing Domains | Periplasmic binding proteins, ligand-binding domains [17] | Analyte recognition and conformational change | Affinity, specificity, conformational dynamics |
| Allosteric TFs | TtgR, TetR, LacI, engineered variants [26] [27] | Ligand-responsive transcriptional regulation | Dynamic range, specificity, orthogonality |
| Linker Sequences | (GGS)n, (EAAAK)n, LEAPAPA [28] | Connect protein domains with controlled flexibility | Length, rigidity, protease resistance |
| Expression Systems | Plasmids, genomic integration sites | Biosensor delivery and maintenance | Copy number, stability, compatibility with host |
| Screening Platforms | Flow cytometry, FACS, microplate readers [29] [27] | High-throughput biosensor characterization | Throughput, sensitivity, multiplexing capability |
The strategic selection of biosensor architecture is paramount for successful investigation of NADPH/NADP+ dynamics in living systems. FRET-based biosensors offer quantitative, ratiometric measurements with high temporal resolution ideal for tracking rapid metabolic fluctuations. Single FP biosensors provide simplified implementation and targeting flexibility, particularly valuable for subcellular compartment-specific monitoring. Transcription factor-based systems enable powerful high-throughput screening applications for strain development and enzyme engineering. Recent advances in computational design, directed evolution, and high-throughput screening methodologies continue to expand the capabilities of each platform, making customized biosensor development more accessible than ever. By applying the appropriate design principles and experimental protocols outlined in this article, researchers can effectively leverage these powerful molecular tools to illuminate the dynamic regulation of cellular redox states.
The dynamic regulation of the NADPH/NADP+ redox couple is central to cellular metabolism, impacting everything from antioxidative defense to reductive biosynthesis. Genetically encoded fluorescent biosensors have revolutionized our ability to monitor these cofactors in living cells with high spatiotemporal resolution. This Application Note provides technical profiles of four leading biosensors—NAPstars, iNAP, mBFP, and NADPsor—framed within the broader research context of dynamically regulating NADPH/NADP+ balance. We detail their working principles, performance characteristics, and provide standardized protocols for their application in metabolic research and drug development.
Technical Biosensor Profiles
The field of NADP(H) biosensing has evolved significantly, with newer sensors offering improved specificity, dynamic range, and practical applicability. The following table summarizes the key characteristics of the featured biosensors.
Table 1: Technical Specifications of Featured NADP(H) Biosensors
| Biosensor Name | Sensing Target | Design Basis | Dynamic Range & Affinity | Key Advantages | Reported Limitations |
|---|---|---|---|---|---|
| NAPstars [3] | NADPH/NADP+ Ratio | Peredox-mCherry chassis with mutated Rex domains | Kr(NADPH/NADP+) from ~0.001 to 5; Kd(NADPH): 0.9 µM (NAPstar1) to 11.6 µM (NAPstar6) [3] | Family of sensors; pH-insensitive; compatible with FLIM [3] | NAPstar6/7 show some pool size dependence [3] |
| iNAP [30] | NADPH | cpYFP inserted into Rex transcriptional repressor | N/A | Bright, ratiometric, large dynamic range [31] | Excitation at short wavelengths; pH-sensitive signal at 480 nm [31] |
| NADPsor [30] | NADP+ | Genetically encoded fluorescent sensor | N/A | Specific for NADP+ [30] | Limited characterization in available literature |
| mBFP | N/A | Note: No technical details were identified in the search results for mBFP. | N/A | N/A | N/A |
| NERNST [32] | NADP(H) Redox Status | roGFP2 fused to NADPH-thioredoxin reductase C (NTRC) | Ratiometric; responds specifically to NADPH, not NADH or GSH [32] | Ratiometric; non-destructive; functional across bacteria, plants, and animals [32] | Specificity concerns due to roGFP2 equilibration with glutathione redox couple [3] |
Detailed Experimental Protocols
Protocol: In Vitro Characterization of NAPstar Biosensors
This protocol describes how to characterize the NADPH dependency and specificity of recombinant NAPstar proteins, as performed in the foundational study [3].
Workflow Overview:
Materials:
- Recombinant NAPstar Protein: Purified from E. coli [3].
- Nucleotides: NADPH, NADP+, NADH, NAD+ (e.g., Sigma-Aldrich).
- Buffer: Suitable physiological buffer (e.g., 50 mM HEPES, pH 7.4).
- Equipment: Spectrofluorometer (e.g., CLARIOstar, BMG LABTECH).
Procedure:
- Protein Purification: Express NAPstar variants (e.g., NAPstar1, 2, 3, 6, 7) in E. coli and purify using affinity chromatography followed by size-exclusion chromatography to isolate dimeric forms [3].
- Spectra Recording: Dilute the purified protein in assay buffer. Record fluorescence excitation and emission spectra at room temperature. For NAPstars, monitor the T-Sapphire (TS) fluorescence (excitation ~400 nm, emission ~515 nm) and the mCherry (mC) signal [3].
- NADPH Titration: Add NADPH to the protein solution in a stepwise manner (e.g., 0.01–1000 µM). At each concentration, measure the TS and mC fluorescence. Calculate the TS/mC emission ratio.
- Specificity Titration: Repeat the titration separately with NADH, NADP+, and NAD+ to determine cross-reactivity.
- Data Analysis:
- Plot the TS/mC ratio against the NADPH concentration.
- Fit the data to determine the apparent dissociation constant for NADPH (Kd(NADPH)).
- To reflect the sensor's response to the NADPH/NADP+ ratio, calculate Kratio (Kr(NADPH/NADP+)) [3].
- To test pool size dependence, perform titrations at different total NADP (NADPH + NADP+) pool sizes (e.g., 100, 300, 500 µM) [3].
Protocol: FACS-Based Enzyme Engineering Using an NADPH Biosensor
This protocol utilizes a transcription factor-based biosensor (pSenSox) for high-throughput screening of NADPH-consuming enzymes, such as alcohol dehydrogenases (ADHs) [33].
Workflow Overview:
Materials:
- Biosensor Plasmid: pSenSox (contains SoxR transcription factor, PsoxS promoter, and eyfp reporter gene) [33].
- Host Strain: E. coli TOP10 or similar [33].
- Library: Mutant library of the NADPH-dependent enzyme of interest (e.g., generated by random mutagenesis of Lactobacillus brevis ADH, LbADH).
- Substrate: The target compound for the enzyme (e.g., 2,5-hexanedione).
- Media: LB or TB medium with appropriate antibiotics [33].
- Equipment: Fluorescence-Activated Cell Sorter (FACS), flow cytometer.
Procedure:
- Library Transformation: Co-transform the mutant enzyme library and the pSenSox biosensor plasmid into E. coli.
- Culture and Induction: Grow transformed cells in a rich medium (e.g., TB). Induce enzyme and biosensor expression with suitable inducers (e.g., IPTG).
- Substrate Challenge: Add the target substrate to the culture. The consumption of NADPH by active enzyme variants triggers the SoxR-based biosensor, leading to eYFP production.
- FACS Analysis and Sorting: Analyze the cell population using FACS. Gate the most fluorescent cells (typically the top 0.1-1%), which correspond to clones with high NADPH consumption and thus high enzyme activity [33].
- Recovery and Validation: Sort the selected population, recover the cells, and isolate the plasmid DNA. Sequence the enzyme gene to identify beneficial mutations. Finally, express and purify the identified variant (e.g., LbADHK71E) for biochemical characterization to confirm improved kinetics [33].
Pathway Diagrams and Logical Workflows
NADP(H) Metabolism and Biosensor Integration
This diagram illustrates the central metabolic pathways involving NADP(H) and the points where different biosensors integrate to monitor its dynamics.
The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential Reagents and Resources for NADP(H) Biosensor Research
| Reagent / Resource | Function/Description | Example Use Case |
|---|---|---|
| pSenSox Plasmid [33] | Transcription factor (SoxR)-based biosensor that activates eYFP expression upon NADPH depletion. | High-throughput screening of mutant libraries for NADPH-consuming enzymes via FACS. |
| pET-TEV Vector [32] | Expression plasmid for producing recombinant biosensor proteins in E. coli. | In vitro characterization and purification of biosensors like NERNST and NAPstars. |
| NADP+, NADPH Standards | High-purity nucleotide cofactors for calibration and in vitro assays. | Generating standard curves for sensor titration and determining Kd/Kr values. |
| FLIM-capable Microscope | Fluorescence Lifetime Imaging Microscope for advanced biosensor readout. | Measuring NAPstar dynamics independent of sensor concentration, using fluorescence lifetime [3]. |
| CLARIOstar Microplate Reader [34] | Versatile microplate reader with UV/VIS spectrometer and fluorescence detection. | Performing ratiometric fluorescence measurements and NAD(P)H conversion assays (e.g., at 340 nm). |
The redox cofactor nicotinamide adenine dinucleotide phosphate (NADPH) is a crucial electron donor for reductive biosynthesis and antioxidative defense in living cells. Maintaining the optimal NADPH/NADP+ ratio is essential for efficient production of valuable chemicals in industrial biotechnology. Traditional static regulation strategies often lead to cofactor imbalance, suboptimal productivity, and cellular stress. This Application Note explores the paradigm shift toward dynamic regulation using genetically encoded biosensors, providing detailed case studies and methodologies for implementing these advanced approaches in strain engineering for biofuel and amino acid production.
Table 1: Key NADPH-Generating Pathways in Microbial Systems
| Pathway | Key Enzymes | Primary Role | NADPH Yield |
|---|---|---|---|
| Oxidative Pentose Phosphate Pathway (oxPPP) | Glucose-6-phosphate dehydrogenase (G6PD), 6-phosphogluconate dehydrogenase (GND) | Major NADPH source in most organisms | Varies by organism and conditions |
| Entner-Doudoroff (ED) Pathway | Glucose-6-phosphate dehydrogenase (G6PD) | Secondary NADPH source, particularly in Pseudomonads | Variable cofactor specificity |
| TCA Cycle | Isocitrate dehydrogenase (IDH) | Supplemental NADPH generation | Organism-dependent |
Static vs. Dynamic Regulation of NADPH
Limitations of Static Regulation Approaches
Static regulation strategies involve permanent genetic modifications that cannot respond to real-time metabolic demands. These include:
- Overexpression of NADPH-generating enzymes (e.g., Zwf, Gnd, IDH)
- Promoter and RBS engineering to enhance pathway flux
- Protein engineering to modify cofactor preference of key enzymes
- Knockout of competing pathways that consume NADPH
While these approaches have successfully improved production of compounds like poly-3-hydroxybutyrate (PHB) and amino acids, they frequently cause NADPH/NADP+ imbalance, leading to disrupted cell growth and suboptimal productivity [35]. The fundamental limitation is their inability to provide real-time adjustment of intracellular NADPH levels in response to changing metabolic demands during different fermentation phases.
Principles of Dynamic Regulation
Dynamic regulation systems utilize genetically encoded components that sense intracellular metabolite levels and respond by modulating gene expression in real-time. These systems typically consist of:
- Biosensors that detect specific metabolites or redox states
- Genetic circuits that process the sensory information
- Actuators that modulate expression of pathway genes
The primary advantage of dynamic regulation is its capacity to maintain redox balance while optimizing carbon flux toward desired products, effectively enabling the microbial chassis to self-optimize its metabolism during fermentation.
Biosensor Platforms for NADPH/NADP+ Monitoring
Available Biosensor Architectures
Recent advances in biosensor development have produced several platforms for monitoring NADPH/NADP+ dynamics:
NAPstar Biosensors: A family of genetically encoded, fluorescent protein-based sensors that specifically measure the NADP redox state (NADPH/NADP+ ratio) across a broad range (0.001 to 5) with subcellular resolution. NAPstars were developed by engineering the NAD redox sensor Peredox-mCherry through mutations that switch specificity from NADH to NADPH [3].
NERNST Biosensor: A ratiometric biosensor that monitors NADP(H) redox status using a redox-sensitive green fluorescent protein (roGFP2) and an NADPH thioredoxin reductase C module, enabling assessment of NADPH/NADP+ balance across organisms [35].
iNap Biosensors: A series of sensors based on the transcription factor Rex that respond to NADPH/NADP+ ratios, with iNap1 demonstrating successful application for real-time monitoring in eukaryotic cells [7].
Table 2: Comparison of NADPH/NADP+ Biosensors
| Biosensor | Detection Principle | Dynamic Range | Compartmentalization | Key Applications |
|---|---|---|---|---|
| NAPstar | FRET-based (Rex domains with cpT-Sapphire) | 0.001 to 5 NADPH/NADP+ ratio | Yes (cytosol, mitochondria) | Real-time monitoring in yeast, plants, mammalian cells |
| NERNST | roGFP2 with TrxR C module | Not specified | Limited data | Cross-organism NADP(H) redox status assessment |
| iNap | Transcription factor-based | Varies by variant | Yes (cytosol, mitochondria) | Live-cell imaging in endothelial cells, high-throughput screening |
Experimental Protocol: Implementing NAPstar Biosensors
Protocol 1: Subcellular NADPH/NADP+ Monitoring with NAPstar Biosensors
Materials:
- NAPstar plasmid DNA (variants 1, 2, 3, 6, or 7 based on desired Kr)
- Appropriate expression system for target organism
- Confocal microscope with capabilities for FRET imaging
- Image analysis software (e.g., ImageJ, MATLAB)
Methodology:
- Sensor Selection: Choose appropriate NAPstar variant based on expected NADPH/NADP+ ratio range:
- NAPstar1: Kr = 0.006 (most reduced)
- NAPstar2: Kr = 0.019
- NAPstar3: Kr = 0.042
- NAPstar6: Kr = 0.077
- NAPstar7: Kr = 0.130 (most oxidized)
Genetic Integration: Transform host organism with NAPstar constructs targeted to specific subcellular compartments (cytosol, mitochondria, etc.) using appropriate molecular biology techniques.
Calibration:
- Perform in situ calibration using digitonin permeabilization (0.001% for plasma membrane, 0.3% for mitochondrial membrane)
- Expose cells to increasing concentrations of NADPH (0-1000 μM) in presence of constant NADP+ (150 μM)
- Generate standard curve of TS/mC fluorescence ratio vs. NADPH/NADP+ ratio
Live-Cell Imaging:
- Culture NAPstar-expressing cells under appropriate conditions
- Acquire time-lapse images using confocal microscopy with excitation at 400/488 nm and emission at 515 nm for TS, and excitation at 587 nm and emission at 610 nm for mCherry
- Calculate ratio of TS/mC fluorescence for each time point
Data Analysis:
- Convert fluorescence ratios to NADPH/NADP+ ratios using calibration curve
- Analyze temporal dynamics and compartment-specific differences
- Correlate NADPH/NADP+ dynamics with metabolic outputs
Troubleshooting:
- If signal-to-noise ratio is low, optimize expression level or try alternative NAPstar variant
- If compartmentalization is unclear, verify targeting sequences and use organelle-specific markers
- For pH sensitivity concerns, monitor intracellular pH concurrently with pH-sensitive dyes
Case Study 1: Biosensor-Driven Isoprenol Production in Pseudomonas putida
Background and Engineering Strategy
Isoprenol is a potential aviation biofuel precursor whose production is often limited by host metabolic constraints. A recent study demonstrated the application of a biosensor-driven selection strategy in Pseudomonas putida to identify and overcome production bottlenecks [36].
The engineering workflow involved:
- Development of an isoprenol-responsive biosensor from native catabolic pathways
- CRISPRi library screening to identify genomic targets that enhance production
- Combinatorial strain engineering based on biosensor selections
- Integrated omics analysis to understand emergent metabolic states
Figure 1: Workflow for biosensor-driven isoprenol production strain development
Experimental Protocol: Biosensor-Driven Strain Selection
Protocol 2: Developing and Implementing Biosensors for Biofuel Production
Materials:
- Bacterial strain (e.g., Pseudomonas putida KT2440)
- Molecular biology reagents for cloning and recombineering
- Fluorescence-activated cell sorting (FACS) capability
- GC-MS for product quantification
Methodology:
- Biosensor Development from Native Systems:
- Identify candidate transcriptional regulators from RB-TnSeq fitness data
- Clone putative promoter regions upstream of fluorescent reporter genes
- Implement RBS optimization to enhance dynamic range
- Characterize dose-response to target molecule (e.g., isoprenol)
Growth-Coupled Selection System:
- Link biosensor activation to essential gene expression
- Implement antibiotic-free selection systems
- Establish correlation between fluorescence and product titer
CRISPRi Library Screening:
- Design gRNA library targeting all open reading frames
- Transform library into production strain with integrated biosensor
- Sort high-fluorescence population using FACS
- Identify enriched gRNAs through whole-genome sequencing
Combinatorial Engineering:
- Select top candidate genes from screening
- Design multiplexed engineering strategies
- Combine targets with rational engineering approaches
- Iterate through multiple engineering cycles
Validation and Analysis:
- Quantify product titers in engineered strains using GC-MS
- Perform transcriptomics and metabolomics on high-performing strains
- Analyze metabolic rewiring and emergent properties
Key Results: The biosensor-driven approach enabled identification of 70 previously untested gene loci and construction of over 165 mutant strains, ultimately yielding a 36-fold increase in isoprenol titer to approximately 900 mg/L. Integrated omics analysis revealed that metabolic rewiring toward amino acid catabolism was crucial for this improvement [36].
Case Study 2: Amino Acid Production in Corynebacterium glutamicum
Systems Metabolic Engineering for L-Lysine
L-Lysine production represents one of the most successful examples of industrial biotechnology, with annual production exceeding 1.5 million tons. Systems metabolic engineering in Corynebacterium glutamicum has demonstrated the critical importance of NADPH supply for high-yield production [37].
Key engineering strategies included:
- Enhancing precursor supply through anaplerotic pathway engineering
- Modulating NADPH generation via pentose phosphate pathway optimization
- Dynamic flux control to balance growth and production phases
- Transport engineering to improve product secretion
Table 3: Metabolic Engineering Targets for Amino Acid Production
| Amino Acid | Host Organism | Key Engineering Targets | NADPH Requirement | Reported Titer |
|---|---|---|---|---|
| L-Lysine | Corynebacterium glutamicum | Dehydrogenase complex, PPP flux, transporter engineering | High (4 NADPH per molecule) | ~120 g/L (industrial processes) |
| L-Threonine | Escherichia coli | Biosynthetic cluster, global regulators, export systems | Moderate | Not specified |
| L-Valine | E. coli and C. glutamicum | Biosynthetic pathway, global regulators, competing pathways | High | 61 g/L in C. glutamicum |
| L-Glutamate | Corynebacterium glutamicum | Anaplerotic reactions, 2-oxoglutarate dehydrogenase | Low | Industrial scale (2.5M tons/year) |
Experimental Protocol: NADPH Optimization for Amino Acid Production
Protocol 3: Balancing NADPH Supply for Amino Acid Biosynthesis
Materials:
- Amino acid production strain (e.g., Corynebacterium glutamicum)
- Enzyme activity assay kits (Zwf, Gnd, IDH)
- Metabolomics platform for NADPH/NADP+ quantification
- Fermentation equipment
Methodology:
- Baseline Characterization:
- Quantify intracellular NADPH/NADP+ ratio across growth phases
- Measure fluxes through NADPH-generating pathways (oxPPP, ED, TCA)
- Determine NADPH demand for target amino acid pathway
- Identify potential cofactor imbalances
Pathway Engineering:
- Overexpress NADPH-generating enzymes (e.g., Zwf, Gnd, IDH)
- Modulate expression strength using promoter/RBS libraries
- Implement protein engineering to alter cofactor specificity
- Knock out competing NADPH-consuming pathways
Dynamic Regulation System:
- Introduce NADPH-responsive biosensor (e.g., NAPstar, iNap)
- Link biosensor to expression of NADPH-generating enzymes
- Implement feedback-controlled fermentation systems
- Optimize induction thresholds for production phase
Fermentation Optimization:
- Develop dual-phase fermentation processes (growth vs. production)
- Implement fed-batch strategies with carbon source control
- Monitor real-time NADPH/NADP+ ratios using online biosensors
- Adjust process parameters based on redox state
Systems Analysis:
- Perform flux balance analysis to predict pathway utilization
- Conduct metabolomics to identify metabolic bottlenecks
- Use transcriptomics to verify regulatory responses
- Integrate multi-omics data for holistic understanding
Key Findings: Successful lysine production strains required global modification of pathway fluxes, with particular emphasis on maintaining optimal NADPH/NADP+ ratios. Engineering the oxidative pentose phosphate pathway and modulating isocitrate dehydrogenase activity proved critical for supplying sufficient reducing power while maintaining redox balance [37].
The Scientist's Toolkit: Essential Research Reagents
Table 4: Key Research Reagents for NADPH Biosensor Studies
| Reagent/Category | Specific Examples | Function/Application | Source/Reference |
|---|---|---|---|
| NADPH Biosensors | NAPstar variants, iNap, NERNST | Real-time monitoring of NADPH/NADP+ ratio | [3] [7] |
| Fluorescent Reporters | mCherry, cpT-Sapphire, roGFP2 | Visualizing biosensor activation and compartmentalization | [3] |
| Enzymatic Assay Kits | G6PDH activity, NADP+ quantification | Validation of biosensor readings | [38] |
| Metabolic Inhibitors | G6PDi-1 (G6PDH inhibitor) | Perturbation of NADPH metabolism for system validation | [39] |
| Analytical Standards | NADP+, NADPH | Calibration of sensors and analytical methods | [38] |
| Molecular Biology Tools | CRISPRi libraries, recombineering systems | High-throughput identification of engineering targets | [36] |
The implementation of dynamic regulation strategies using advanced biosensors represents a paradigm shift in metabolic engineering for biofuel and amino acid production. The case studies presented demonstrate that biosensor-driven approaches can successfully overcome the limitations of traditional static engineering, enabling unprecedented improvements in product titers.
Future developments in this field will likely focus on:
- Multiparameter biosensing for simultaneous monitoring of multiple cofactors and metabolites
- Machine learning integration for predictive modeling of metabolic dynamics
- Closed-loop bioprocessing with real-time control based on biosensor inputs
- Cross-kingdom application of biosensor technologies from microbes to mammalian systems
The continued refinement of these tools and strategies will accelerate the development of efficient microbial cell factories for sustainable production of biofuels, biochemicals, and therapeutic compounds.
The dynamic regulation of the NADPH/NADP+ redox couple is central to metabolic homeostasis and cellular responses to stress across diverse biological systems. The development of genetically encoded biosensors has revolutionized our ability to monitor these dynamics in real-time within living cells. This Application Notes and Protocols document provides a detailed guide for implementing the latest NADPH/NADP+ biosensing technologies, specifically the NAPstar family of biosensors, in yeast, mammalian cells, and plant models. The protocols are framed within the broader thesis of understanding how cells maintain redox balance across different compartments and organisms, with particular emphasis on applications in dynamic environmental conditions including illumination-dependent responses in plants.
NAPstar Biosensor Family
The NAPstar biosensor family represents a significant advancement in redox metabolism monitoring. These genetically encoded, fluorescent protein-based biosensors specifically measure the NADPH/NADP+ redox state with subcellular resolution across a broad dynamic range [3] [40]. Derived from the bacterial transcriptional repressor Rex, NAPstars incorporate a circularly permuted T-Sapphire (TS) fluorescent protein nested between two NADH/NAD+-binding domains that have been engineered for NADP specificity through rational mutagenesis [3].
Key advantages of NAPstars include:
- Specificity: High specificity for NADPH/NADP+ over NADH/NAD+ redox couples
- pH stability: Limited sensitivity to pH fluctuations
- Dual readout: Ratiometric measurements via TS/mCherry fluorescence
- Versatility: Compatibility with both fluorescence intensity and fluorescence lifetime imaging (FLIM)
- Broad range: Capacity to monitor NADPH/NADP+ ratios across a 5000-fold range (approximately 0.001 to 5) [3]
Table 1: Characteristics of Selected NAPstar Biosensor Variants
| Biosensor Variant | Kd(NADPH) (μM) | Kratio(NADPH/NADP+) | Dynamic Range | Preferred Applications |
|---|---|---|---|---|
| NAPstar1 | 0.9 | 0.006 | ~2.5 | Cytosolic measurements |
| NAPstar3 | 2.2 | 0.015 | ~2.5 | General purpose |
| NAPstar6 | 11.6 | 0.077 | ~2.5 | High-ratio environments |
| NAPstarC | No binding | No response | None | Negative control |
Research Reagent Solutions
Table 2: Essential Materials for NADPH/NADP+ Biosensor Research
| Reagent/Material | Function/Application | Examples/Specifications |
|---|---|---|
| NAPstar Biosensors | Genetically encoded redox state monitoring | NAPstar1-7, NAPstarC (control) |
| Plasmid Vectors | Biosensor delivery to cells | Mammalian: pcDNA3.1, Plant: pBIN19, Yeast: YEplac series |
| Cell Lines | Experimental model systems | HeLa (mammalian), Saccharomyces cerevisiae (yeast), Arabidopsis thaliana (plant) |
| Microscopy Systems | Biosensor signal detection | Confocal microscopy with 400/515 nm (TS) and 587/610 nm (mCherry) filters |
| FLIM Capability | Fluorescence lifetime measurement | Alternative readout method for biosensor signal |
| Two-Photon Microscopy | Deep tissue imaging in plants | Minimizes chlorophyll autofluorescence [41] |
| Dynamic Microfluidics | Environmental perturbation studies | dMSCC for rapid media switching [42] |
Application Protocols
Protocol 1: Monitoring NADP Redox Dynamics in Yeast During Cell Division
Background: Yeast cells exhibit oscillations in NADP redox state correlated with metabolic cycles and cell division. This protocol enables real-time monitoring of these dynamics using NAPstar biosensors [3].
Materials:
- Saccharomyces cerevisiae strains (e.g., CEN.PK113-7D, Ethanol Red, PE2) [42]
- NAPstar expression vector (e.g., NAPstar3 for general purpose)
- Standard yeast growth media (YPD or synthetic complete)
- Microfluidic cultivation system (dMSCC) [42]
- Confocal or fluorescence microscopy system with environmental control
Procedure:
- Transform yeast strains with NAPstar plasmids using standard lithium acetate transformation.
- Select positive clones on appropriate selective media and verify expression.
- Calibrate the biosensor in vivo using established calibration protocols [3].
- Culture transformed yeast in microfluidic chambers under controlled conditions.
- Implement dynamic perturbations using dMSCC to simulate industrial bioreactor conditions:
- Image cells every 2-5 minutes using time-lapse microscopy.
- Quantify fluorescence ratios (TS/mCherry) and convert to NADPH/NADP+ ratios.
- Correlate redox oscillations with cell cycle stage (budding index).
Expected Results: Yeast typically show regular oscillations in NADPH/NADP+ ratio during cell division, with strain-specific variations in robustness to substrate oscillations [3] [42].
Protocol 2: Assessing Oxidative Stress Responses in Mammalian Cells
Background: The glutathione system plays a conserved, primary role in mediating antioxidative electron flux across eukaryotes. This protocol uses NAPstars to quantify NADP redox changes during oxidative challenge in mammalian cells [3] [40].
Materials:
- Mammalian cell lines (HeLa, HEK293, or other relevant models)
- NAPstar expression vectors (NAPstar1 recommended for cytosolic measurements)
- Transfection reagent (e.g., Lipofectamine 3000)
- Oxidative stress inducers: H₂O₂, menadione, or diamide
- Inhibitors: Buthionine sulfoximine (BSO) for glutathione depletion
- Live-cell imaging chamber with environmental control (37°C, 5% CO₂)
Procedure:
- Culture mammalian cells in appropriate media (DMEM for HeLa) on glass-bottom dishes.
- Transfect with NAPstar plasmids using standard transfection protocols.
- Incubate for 24-48 hours to allow biosensor expression.
- Pre-treat cells with pathway-specific inhibitors:
- Glutathione pathway: 100 μM BSO for 4 hours
- Thioredoxin pathway: Selective inhibitors as available
- Mount dishes on live-cell imaging system with temperature and CO₂ control.
- Acquire baseline images for 5-10 minutes before stress induction.
- Induce oxidative stress with 100-500 μM H₂O₂.
- Image continuously for 60-90 minutes post-induction.
- Quantify fluorescence ratios and calculate NADPH/NADP+ ratios.
- Compare kinetics between control and inhibitor-treated cells.
Expected Results: The glutathione system typically demonstrates primacy in NADPH-dependent antioxidative electron flux, with faster and more pronounced changes in NADP redox state when this pathway is impaired [3] [40] [43].
Protocol 3: Illumination-Dependent NADP Redox Monitoring in Plants
Background: Plant chloroplasts undergo rapid NADP redox state changes in response to light availability. This protocol details the use of NAPstars or alternative biosensors (mBFP) to monitor these dynamics [3] [41].
Materials:
- Arabidopsis thaliana or Nicotiana benthamiana plants
- NAPstar vectors with chloroplast targeting or Tha4-f:mt-mBFP for thylakoid targeting [41]
- Agrobacterium tumefaciens for plant transformation (floral dip or infiltration)
- Two-photon microscopy system to minimize chlorophyll autofluorescence [41]
- Controlled light source with adjustable wavelength and intensity
Procedure:
- Generate transgenic plants expressing chloroplast-targeted NAPstars or:
- For transient expression: Infiltrate N. benthamiana leaves with Agrobacterium carrying biosensor constructs
- For mBFP-based sensors: Use Tha4-f:mt-mBFP targeted to thylakoid membrane [41]
- Mount leaf samples under two-photon microscope with controlled light source.
- Optimize detection: Use 720-760 nm excitation for mBFP to minimize chlorophyll interference [41]
- Acquire baseline measurements in dark-adapted plants (30 minutes minimum).
- Apply light stimuli with varying:
- Intensities: 50-1000 μmol photons m⁻² s⁻¹
- Wavelengths: Blue (400-500 nm), red (600-700 nm), or full spectrum
- Image continuously during light-dark transitions.
- For hypoxia studies: Apply nitrogen atmosphere to create hypoxic conditions.
- Quantify fluorescence changes and convert to relative NADPH levels or NADPH/NADP+ ratios.
Expected Results: Rapid increase in stromal NADPH upon illumination, with wavelength-dependent kinetics. Stronger responses typically to blue light due to photoreceptor activation [3] [41] [44].
Data Analysis and Interpretation
Quantitative Analysis of NADP Redox State
Calibration Procedures: For accurate quantification of NADPH/NADP+ ratios, in vivo calibration is essential:
For NAPstars: Use the established relationship between TS/mCherry fluorescence ratio and NADPH/NADP+ ratio:
Where R is the measured TS/mCherry ratio, Roxidized is the ratio in fully oxidized state, and Rreduced is the ratio in fully reduced state.
Determine in vivo extremes:
- Fully oxidized state: Treat with 10 mM diamide for 10 minutes
- Fully reduced state: Treat with 10 mM dithiothreitol (DTT) for 10 minutes
Calculate NADPH/NADP+ ratio using the sensor-specific Kratio values provided in Table 1.
Statistical Considerations:
- Perform experiments with at least 3-5 biological replicates
- Analyze minimum 20-30 cells per condition for single-cell analyses
- Account for cell-to-cell heterogeneity in population responses
Troubleshooting Common Issues
Table 3: Troubleshooting Guide for NADPH/NADP+ Biosensing
| Problem | Potential Cause | Solution |
|---|---|---|
| Low signal-to-noise ratio | Poor biosensor expression | Optimize transfection/transformation; check promoter strength |
| Unresponsive biosensor | Incorrect subcellular targeting | Verify targeting sequences; confirm localization |
| Photobleaching | Excessive illumination intensity | Reduce excitation intensity; use two-photon microscopy |
| Chlorophyll interference (plants) | Standard fluorescence detection | Switch to two-photon microscopy [41] |
| Non-specific oxidative changes | Sensor cross-talk with glutathione pool | Use NAPstars instead of roGFP-based sensors [3] |
The protocols outlined herein provide researchers with robust methodologies for investigating NADPH/NADP+ redox dynamics across yeast, mammalian cells, and plant systems. The application of these biosensors has already revealed fundamental biological insights, including the conserved robustness of cytosolic NADP redox homeostasis, cell cycle-linked oscillations in yeast, and the primacy of the glutathione system in antioxidative defense across eukaryotes [3] [40] [43].
Future applications of these technologies may include:
- High-throughput screening for redox-modulating compounds in drug discovery
- Optimization of industrial microbial strains for bioproduction
- Development of stress-resilient crop varieties through redox profiling
- Integration with other biosensors for multi-parameter metabolic monitoring
As the field advances, these protocols will enable deeper investigation into the fundamental principles governing cellular redox regulation and its applications in biotechnology and medicine.
Overcoming Hurdles: A Troubleshooting Guide for Biosensor Implementation and Optimization
The accurate measurement of the NADPH/NADP+ redox couple is fundamental to understanding cellular redox metabolism, anabolic processes, and antioxidant defense [3] [30]. However, the close structural similarity between NAD(P)H and the presence of other cellular redox systems, particularly the glutathione (GSH/GSSG) couple, presents a significant challenge for specific measurement [3] [30]. The tripeptide glutathione is a ubiquitous cellular antioxidant that maintains redox homeostasis and detoxifies reactive oxygen species, often operating in concert with NADPH-dependent enzymes like glutathione reductase [45]. Cross-reactivity and interference from these systems can compromise data integrity, leading to inaccurate assessment of the NADP redox state. This Application Note details strategies and protocols to minimize such cross-reactivity, ensuring robust and specific NADP(H) measurements within the context of dynamic NADPH/NADP+ balance regulation research. The recommendations are crucial for researchers employing biosensors to investigate metabolic engineering, drug mechanisms, or redox biology in live cells.
Biosensor Engineering for Intrinsic Specificity
The most fundamental strategy for ensuring specificity begins with the rational design of the biosensor protein itself. Exploiting the structural differences between NADPH and its close analog NADH through targeted mutagenesis has proven highly successful.
The NAPstar Family of Biosensors
The NAPstar biosensor family, developed from the Peredox-mCherry chassis, exemplifies this approach [3]. Peredox was originally designed to sense the NADH/NAD+ ratio using the NADH-binding domain of the bacterial Rex transcription factor. To achieve NADP(H) specificity, researchers introduced specific mutations known to switch the binding pocket's preference from NADH to NADPH [3]. The resulting NAPstar sensors demonstrate a pronounced affinity for NADPH over NADH, with apparent dissociation constants (Kd(NADPH)) ranging from 0.9 to 11.6 µM, while their affinity for NADH was one to two orders of magnitude lower (Kd(NADH) from 24.4 to 248.9 µM) [3]. This dramatic shift in specificity is a direct result of strategic protein engineering.
Table 1: Characterization of Select NAPstar Sensor Variants
| Sensor Variant | Kd(NADPH) (µM) | Kd(NADH) (µM) | Specificity Ratio (Kd(NADH)/Kd(NADPH)) | Key Property |
|---|---|---|---|---|
| NAPstar1 | 0.9 | 24.4 | ~27 | Highest NADPH affinity |
| NAPstar3 | 2.6 | 43.4 | ~17 | Balanced affinity and dynamic range |
| NAPstar6 | 11.6 | 248.9 | ~21 | Lower affinity, wider dynamic range |
Selectivity Against Glutathione Systems
A critical validation step for any NADP(H) biosensor is testing against the glutathione redox couple. The NERNST sensor, which relies on a roGFP2 reporter coupled to NADPH-thioredoxin reductase C, was found to efficiently equilibrate with the glutathione pool in vivo, confounding measurements [3]. In contrast, the NAPstar family functions independently of the glutathione and thioredoxin systems, as confirmed through experiments in yeast, plant, and mammalian cells where these antioxidative pathways were genetically impaired [3]. This orthogonality is essential for attributing observed redox dynamics specifically to the NADP pool.
Experimental and Measurement Strategies to Minimize Interference
Even with a specific biosensor, experimental design and appropriate controls are vital to account for potential residual interference or context-specific factors.
Ratiometric Measurement and Control Sensors
A primary method to control for non-specific environmental effects is the use of ratiometric biosensors. For example, NAPstars and Peredox incorporate a C-terminal mCherry (mC) fluorescent protein. The NAD(P)H-sensitive fluorescent protein (e.g., T-Sapphire) emission is normalized against the mCherry signal, which is inert to the redox state [3] [46]. This ratio (TS/mC) corrects for variations in sensor expression, cell thickness, and photobleaching.
Furthermore, employing a non-binding control sensor is crucial. NAPstarC, which contains mutations that prevent nucleotide binding, serves as a perfect negative control. Its signal should remain unresponsive to changes in NADPH concentration, and any observed fluctuations can be attributed to non-specific experimental artifacts [3].
Sentinel Sensors and Permselective Membranes
For electrochemical biosensors, a common strategy is to use a "sentinel" sensor, which is identical to the biosensor but lacks the biorecognition element (e.g., the Rex domain) or contains an inactivated enzyme [47]. The signal from this sentinel sensor, which records only the background and interference currents, can be subtracted from the biosensor's total signal to yield a specific response.
The use of permselective membranes, such as Nafion or cellulose acetate, can also enhance selectivity. These membranes can be coated over the sensor to prevent interfering anionic species (e.g., ascorbate, urate) or larger molecules from reaching the electrode surface based on charge or size exclusion [47].
Pharmacological and Genetic Validation
Confirming biosensor readings through independent methods is a cornerstone of validation.
- Pharmacological Inhibition: Using specific inhibitors of key NADPH-consuming or producing pathways (e.g., the glutathione system) can help verify the source of the signal. If the biosensor is specific, its signal should not directly correlate with perturbations that only affect the glutathione pool.
- Genetic Manipulation: Knocking out or overexpressing genes involved in NADPH metabolism (e.g., glucose-6-phosphate dehydrogenase) and observing the corresponding biosensor response provides a powerful validation of its specificity [3]. Similarly, as performed with NAPstars, impairing the glutathione and thioredoxin pathways demonstrates that the sensor remains functional and reports specifically on NADP(H) dynamics even when these parallel systems are compromised [3].
Protocol: Validating NADP(H) Biosensor Specificity
This protocol outlines the key steps for establishing the specificity of an NADP(H) biosensor, such as a NAPstar variant, in a live-cell imaging experiment.
In Vitro Characterization of Sensor Specificity
Purpose: To determine the affinity and cross-reactivity of the biosensor for NADPH, NADH, NADP+, and NAD+ in a controlled, cell-free system. Materials:
- Purified biosensor protein (e.g., NAPstar)
- Control sensor (e.g., NAPstarC)
- NADPH, NADH, NADP+, NAD+ (prepare fresh stock solutions)
- Assay buffer (e.g., 50 mM Tris-HCl, 100 mM NaCl, pH 7.4)
- Fluorescence plate reader or spectrophotometer capable of ratiometric measurements (e.g., excitation at 400/485 nm, emission at 515/610 nm)
Procedure:
- Dilute the purified biosensor to a consistent concentration in assay buffer.
- For NADPH/NADH titration: In a series of cuvettes or plate wells, maintain a constant, saturating concentration of NADP+ (e.g., 150 µM) and titrate NADPH from 0 to 1000 µM. Repeat for NADH in the presence of constant NAD+ (e.g., 500 µM).
- For NADP+/NAD+ titration: Titrate NADP+ or NAD+ in the absence of reduced equivalents to observe the reverse response.
- After each addition, mix thoroughly and record the fluorescence emission ratios (e.g., TS/mC).
- Plot the ratiometric signal against the concentration of the titrated nucleotide.
- Fit the data to a binding isotherm (e.g., Hill equation) to calculate the apparent Kd for each nucleotide.
Expected Outcome: A specific NADP(H) biosensor will show a strong, saturable response to NADPH and NADP+, but a minimal response to NADH and NAD+ over physiologically relevant concentration ranges, as seen in Table 1.
In Cellulo Validation Using Sentinel and Control Sensors
Purpose: To control for cellular autofluorescence, environmental effects, and non-specific binding in live cells. Materials:
- Cells expressing the NADP(H) biosensor (e.g., NAPstar3)
- Cells expressing a non-binding control sensor (e.g., NAPstarC)
- Cells expressing a sentinel sensor (e.g., mCherry alone)
- Confocal or fluorescence microscope with ratiometric capabilities
- Imaging chamber with controlled environment (CO2, temperature)
Procedure:
- Culture and seed cells expressing the different sensors in an imaging-compatible chamber.
- Acquire baseline ratiometric images for all cell lines using identical acquisition settings.
- Apply a stimulus known to alter NADPH levels (e.g., oxidative challenge with H2O2, nutrient shift).
- Monitor and record the ratiometric changes over time.
- Data Analysis: Subtract the signal from the sentinel sensor (mCherry-only) from the biosensor's mCherry channel if needed for bleed-through correction. The signal from the non-binding control (NAPstarC) should remain constant; any significant change indicates a non-specific artifact. True NADP(H)-specific signals are those observed only in the functional biosensor-expressing cells and are absent in the control cells.
The Scientist's Toolkit: Key Reagents for Specific NADP(H) Sensing
Table 2: Essential Research Reagents and Their Applications
| Reagent / Tool | Function / Description | Key Use Case |
|---|---|---|
| NAPstar Biosensors | A family of genetically encoded, ratiometric biosensors with high specificity for the NADPH/NADP+ ratio. | Real-time, subcellular monitoring of NADP redox state in live yeast, plant, and mammalian cells [3]. |
| Peredox | A genetically encoded, ratiometric biosensor for the NADH/NAD+ ratio. | Serves as a chassis for NAPstar development; useful for parallel monitoring of the NAD pool [3] [46]. |
| NAPstarC | A non-binding control version of the NAPstar sensor. | Critical control for identifying non-specific signal changes due to pH, temperature, or other cellular artifacts [3]. |
| SoxR Biosensor | A transcription factor-based biosensor that responds to the NADPH/NADP+ ratio. | Useful for dynamic regulation of gene expression based on NADPH status in E. coli [35]. |
| roGFP-based Sensors | Redox-sensitive GFP sensors that equilibrate with glutaredoxin and report on the GSH/GSSG redox potential. | Important Note: Can cross-react with glutathione system; use to monitor the interfering pool, not for specific NADPH measurement [3] [35]. |
| Permselective Membranes | Membranes like Nafion or cellulose acetate that block charged interferents. | Coating for electrochemical biosensors to prevent access of ascorbate and urate to the electrode surface [47]. |
Visualizing Strategies and Workflows
Specificity Validation Workflow
The following diagram illustrates the multi-faceted approach to ensuring biosensor specificity, from initial design to final validation.
Relationship Between Redox Systems
This diagram clarifies the relationship and potential points of cross-talk between the NADPH and glutathione redox systems that specificity strategies must overcome.
The dynamic regulation of the NADPH/NADP+ redox couple is fundamental to cellular metabolism, redox signaling, and antioxidative defense. Genetically encoded biosensors have revolutionized our ability to monitor these dynamics with subcellular resolution in living systems. The performance of these biosensors is not inherent but is critically dependent on three optimization parameters: linker lengths that control structural flexibility, binding pocket engineering that dictates metabolite specificity, and expression levels that determine signal fidelity. This Application Note provides a structured framework for optimizing these parameters, with a specific focus on the recently developed NAPstar family of NADP redox state biosensors, to ensure robust measurement of NADPH/NADP+ balance in diverse experimental contexts [3].
Biosensor Optimization Parameters
Quantitative Performance of NAPstar Biosensor Variants
The NAPstar biosensor family, derived from the bacterial transcriptional repressor Rex, offers a range of variants with differing affinities for NADPH and NADP+, enabling researchers to select the optimal tool for their specific experimental system and expected metabolite concentrations [3].
Table 1: Characterization of NAPstar Biosensor Variants
| Biosensor Variant | Kd(NADPH) (μM) | Kr(NADPH/NADP+) | Dynamic Range | Recommended Applications |
|---|---|---|---|---|
| NAPstar1 | 0.9 | - | ~2.5-fold | High-affinity detection in low NADPH environments |
| NAPstar2 | - | - | ~2.5-fold | General purpose |
| NAPstar3 | - | - | ~2.5-fold | General purpose, balanced affinity |
| NAPstar6 | 11.6 | - | ~2.5-fold | Lower affinity for high-concentration dynamics |
| NAPstar7 | - | - | ~2.5-fold | Lower affinity for high-concentration dynamics |
| NAPstarC | Non-binding | Non-responsive | - | Negative control for expression and background |
Research Reagent Solutions
A successful biosensing experiment relies on the availability of specific genetic tools and reagents. The table below lists key materials for implementing NAPstar-based NADPH/NADP+ sensing.
Table 2: Essential Research Reagents for NADP Redox State Monitoring
| Reagent / Tool Name | Type/Function | Key Features and Applications |
|---|---|---|
| NAPstar Plasmid Family | Genetically encoded biosensor | A family of variants (e.g., NAPstar1, 3, 6, 7) with different NADPH affinities for flexible application [3]. |
| pSenSox Biosensor | Transcription factor-based NADPH biosensor | Used in E. coli for high-throughput screening of NADPH-consuming enzymes via FACS [48]. |
| NADP-Snifit | Semisynthetic biosensor | Ratiometric, pH-insensitive sensor for NADPH/NADP+ ratios; excited at long wavelengths (560 nm) [31]. |
| pcDNA3.1(+) NAPstar3b | Mammalian expression vector | Plasmid for expressing the NAPstar3b biosensor in mammalian cell lines [49]. |
| p413 TEF NAPstar Series | Yeast expression vectors | Plasmids for expressing various NAPstar biosensors in S. cerevisiae under the TEF promoter [49]. |
Experimental Protocols
Protocol 1: Optimizing Biosensor Expression Levels in Mammalian Cells
Principle: Excessive biosensor expression can buffer the target metabolite, altering native concentrations and dynamics. This protocol establishes a workflow to titrate expression to reporting levels.
Materials:
- Mammalian expression vector (e.g., pcDNA3.1(+) NAPstar3b [49])
- Cell culture reagents and transfection reagent
- Fluorescence microscope or flow cytometer
Procedure:
- Transfection Titration: Transfert cells with a range of plasmid DNA concentrations (e.g., 0.5, 1.0, 2.0 µg/well in a 6-well plate) using a standard transfection protocol.
- Expression Monitoring: 24-48 hours post-transfection, image cells or analyze by flow cytometry to measure the mCherry reference signal (excitation ~587 nm, emission ~610 nm).
- Functional Validation: Select the lowest expression level that yields a robust and measurable ratiometric signal (TS/mCherry) upon application of a positive control (e.g., oxidative stress inducer).
- Metabolite Buffering Control: Compare the measured NADP redox state in cells with high versus low biosensor expression. A significant difference indicates buffering, and the lower expression level should be selected for experiments.
Protocol 2: In Vitro Characterization of Binding Affinity and Dynamic Range
Principle: Determining the dissociation constant (Kd) and dynamic range of a biosensor variant is crucial for selecting the appropriate tool and interpreting in vivo data.
Materials:
- Purified NAPstar protein (e.g., NAPstar1, 3, 6) [3]
- Fluorescence plate reader capable of dual-excitation ratiometry
- NADPH, NADP+, NADH, NAD+ stocks
Procedure:
- Sample Preparation: Dilute purified biosensor to a working concentration in a physiologically relevant buffer (e.g., pH 7.4).
- NADPH Titration: Aliquot the biosensor into a 96-well plate. Titrate with NADPH (e.g., 0.01–1000 µM) while keeping NADP+ concentration constant (e.g., 150 µM).
- Fluorescence Measurement: For each titration point, measure the fluorescence intensity with excitation at ~400 nm (TS) and ~587 nm (mCherry). Calculate the TS/mCherry ratio.
- Data Analysis: Plot the ratiometric change against the NADPH concentration. Fit the data with a non-linear regression (saturation binding curve) to determine the Kd(NADPH).
- Specificity Testing: Repeat the titration with NADH in the presence of NAD+ to confirm specificity for the phosphorylated cofactor.
Protocol 3: FACS-Based Screening for Enzyme Engineering
Principle: The pSenSox biosensor responds to NADPH consumption by activating a fluorescent reporter, enabling high-throughput isolation of efficient enzyme variants via Fluorescence-Activated Cell Sorting (FACS) [48].
Materials:
- pSenSox biosensor plasmid in E. coli [48]
- Library of enzyme variants (e.g., LbADH mutant library)
- FACS instrument
- Target substrate (e.g., 2,5-hexanedione)
Procedure:
- Library Transformation: Transform the pSenSox plasmid, carrying the mutant enzyme library, into an appropriate E. coli host strain.
- Induction and Substrate Exposure: Grow the cell library to mid-log phase, induce enzyme expression, and add the target substrate to initiate the NADPH-consuming reaction.
- FACS Sorting: Sort the cell population based on the biosensor's reporter fluorescence (e.g., eYFP). Cells with lower fluorescence have consumed more NADPH, indicating higher enzyme activity.
- Validation and Sequencing: Collect the sorted population, plate for single colonies, and reassay individual clones for activity. Sequence validated hits to identify beneficial mutations.
Visualization of Workflows and Mechanisms
NAPstar Biosensor Mechanism and Workflow
FACS Screening with pSenSox Biosensor
The precise optimization of linker design, binding pocket affinity, and expression levels is paramount for leveraging the full potential of genetically encoded biosensors in NADPH/NADP+ research. The protocols and data outlined herein provide a standardized approach for maximizing the signal-to-noise ratio and biological relevance of measurements obtained with the NAPstar family and related biosensors. By systematically applying these optimization strategies, researchers can reliably uncover the dynamic regulation of central redox metabolism in health, disease, and throughout the drug discovery process.
The dynamic regulation of the NADPH/NADP+ balance is fundamental to cellular redox biology, influencing processes from anabolic metabolism to antioxidative defense and cellular signaling. Genetically encoded biosensors have revolutionized our ability to monitor these dynamics in living systems with subcellular resolution. The NAPstar family of biosensors, derived from the Peredox-mCherry chassis but engineered for NADP(H) specificity, represents a significant advancement in the field [3]. These sensors incorporate a circularly permuted T-Sapphire fluorescent protein nested between two bacterial Rex NADH/NAD+-binding domains that have been mutated to favor NADPH binding [3]. Despite these improvements, critical technical challenges persist in their application, primarily concerning pool size dependence, pH sensitivity, and the complexities of in vivo calibration. This application note details these challenges and provides standardized protocols to enhance the reliability and interpretation of NADPH/NADP+ biosensing data, framed within the broader context of optimizing redox balance regulation.
Table 1: Key Characteristics of Representative NADP(H) Biosensors
| Biosensor Name | Base Scaffold | Reported Dynamic Range (NADPH/NADP+) | Key Advantages | Primary Limitations |
|---|---|---|---|---|
| NAPstar Family [3] | Peredox-mCherry (Rex domains) | 0.001 to 5 | Limited pH sensitivity; High brightness; Compatible with FLIM | Variant-specific pool size dependence (NAPstar6/7) |
| iNap Sensors [3] | cpYFP-based | Not specified in context | High specificity for NADPH | Lower brightness; Sensitivity to pH changes |
| NERNST [3] | roGFP2 | Not specified in context | Ratiometric measurement | Potential equilibration with glutathione pool |
Technical Challenge 1: Pool Size Dependence
Understanding the Problem
Pool size dependence refers to a biosensor's response being influenced not only by the NADPH/NADP+ ratio but also by fluctuations in the total concentration of the NADP pool (NADPH + NADP+). This confounds data interpretation, as observed signal changes could stem from altered redox state, altered total metabolite abundance, or both. Investigations with the NAPstar family revealed that while sensors like NAPstar1-3 maintain stable responses across varying total NADP pool sizes (100, 300, and 500 µM), variants with higher dissociation constants, such as NAPstar6 and NAPstar7, exhibit significant pool size dependence [3]. This suggests that sensors with lower affinity for NADPH are more susceptible to this artifact, as their binding sites are not saturated under physiological concentrations.
Experimental Protocol: Validating Pool Size Independence
Objective: To characterize and control for the pool size dependence of a NADPH/NADP+ biosensor in vitro.
Materials:
- Purified biosensor protein (e.g., NAPstar variant)
- NADPH stock solution
- NADP+ stock solution
- Assay buffer (e.g., 50 mM HEPES, 100 mM KCl, pH 7.4)
- Fluorimeter or plate reader capable of time-based measurements
Procedure:
- Prepare NADP+ Master Solutions: Create three separate master solutions of NADP+ in assay buffer with final concentrations of 100 µM, 300 µM, and 500 µM.
- Titrate with NADPH: For each master solution, prepare a dilution series where the NADPH concentration varies (e.g., from 0.01 µM to 1000 µM) while the NADP+ concentration is held constant.
- Measure Sensor Response: Add a fixed concentration of the purified biosensor to each condition and record the fluorescence signal (e.g., T-Sapphire/mCherry ratio for NAPstars).
- Data Analysis: Plot the sensor's response (e.g., fluorescence ratio) against the NADPH/NADP+ ratio. Overlay the curves obtained from the three different total pool sizes. A pool-independent sensor will show superimposable curves, while a dependent sensor will show shifted curves.
Interpretation and Mitigation:
- Sensor Selection: Prioritize sensors like NAPstar1, 2, or 3 that demonstrate minimal pool size dependence across a physiologically relevant range [3].
- Contextual Interpretation: When using sensors with known dependence (e.g., NAPstar6/7), avoid interpreting ratio changes in isolation. Corroborate findings with other methods that report on the total pool size or the absolute concentration of NADPH.
Technical Challenge 2: pH Sensitivity
Understanding the Problem
Cellular pH can fluctuate significantly between compartments and in response to metabolic states. Since the fluorescence of many engineered biosensors is inherently sensitive to their ionic environment, pH changes can masquerade as changes in the target metabolite, leading to false conclusions. The NAPstar family was specifically developed to minimize this confounder, benefiting from the limited pH sensitivity of the Peredox chassis [3]. However, this challenge is starkly illustrated by other sensor types, such as those based on polyaniline (PANI), where the protonation state of the polymer is the fundamental mechanism of sensing, making them intrinsically pH-sensitive [50]. Furthermore, the intrinsic fluorescence of NAD(P)H itself, often used in FLIM, is also sensitive to its biochemical environment and binding to enzymes, which can be influenced by pH [51].
Experimental Protocol: Assessing pH Sensitivity
Objective: To determine the pH sensitivity of a NADPH/NADP+ biosensor and establish a correction factor.
Materials:
- Purified biosensor protein
- A series of pH-buffered solutions (e.g., pH 6.5, 7.0, 7.5, 8.0) with constant ionic strength
- Fluorimeter or plate reader
Procedure:
- Calibrate at Fixed Redox State: Prepare a solution containing the biosensor and a fixed, known NADPH/NADP+ ratio.
- Measure Across pH Range: Divide this solution into aliquots and adjust each to a specific pH within the physiological range using the different buffered solutions. Measure the fluorescence output (intensity or ratio) at each pH value.
- Generate Correction Curve: Plot the fluorescence signal against the pH to create a standard curve. This curve quantifies the pH-dependent change in signal independent of the redox state.
Interpretation and Mitigation:
- In Situ Calibration: For in vivo experiments, use ionophores (e.g., nigericin) in high-K+ buffers to clamp intracellular pH at known values and build a cell-specific correction curve.
- Ratiometric Design: Leverage the built-in calibration of ratiometric sensors like NAPstars, where the pH-sensitive reporter (T-Sapphire) is normalized against a pH-insensitive reference (mCherry) [3].
- Parallel Monitoring: Whenever possible, concurrently monitor intracellular pH using a dedicated pH biosensor to deconvolve its contribution from the NADPH/NADP+ signal.
Technical Challenge 3: In Vivo Calibration & Validation
Understanding the Problem
Translating a biosensor's fluorescence readout into a physiologically meaningful NADPH/NADP+ ratio within the complex environment of a living cell is the ultimate challenge. It requires strategies to clamp or manipulate the redox state to known extremes (fully oxidized and fully reduced) without compromising cell viability. Furthermore, validating sensor readings against orthogonal methods is crucial for establishing credibility. Recent work on electrochemical DNA (eDNA) biosensors highlights the field's drive toward "calibration-free" operational strategies for in vivo monitoring, though this remains an area of active development [52].
Experimental Protocol: Two-Point In Vivo Calibration
Objective: To perform an in vivo calibration of a NADPH/NADP+ biosensor in a cell culture model.
Materials:
- Cell line stably expressing the NADPH/NADP+ biosensor (e.g., NAPstar)
- Imaging chamber with controlled environment (CO₂, temperature)
- Confocal or fluorescence microscope
- Calibration reagents:
- Oxidizing Agent: Hydrogen peroxide (H₂O₂, e.g., 1-10 mM)
- Reducing Agent: Dithiothreitol (DTT, e.g., 1-5 mM) or similar
- Inhibitor: Rotenone (e.g., 1-5 µM) to inhibit mitochondrial ETC and maximize reduction
Procedure:
- Baseline Recording: Image cells in standard culture medium to establish the baseline fluorescence ratio (Rₘᵢₙ).
- Induce Oxidized State (Rₘᵢₙ): Treat cells with a bolus of H₂O₂ to oxidize the NADP pool. The ratio should stabilize at a minimum value. Confirm with subsequent application of rotenone, which should not increase the ratio.
- Induce Reduced State (Rₘₐₓ): Wash out oxidants and treat cells with a combination of rotenone (to block oxidation) and DTT (to chemically reduce the pool). The ratio should reach a maximum value.
- Calculate Ratio: The measured ratio (R) in untreated or experimental conditions can be converted to the NADPH/NADP+ ratio using the formula: NADPH/NADP+ = (R - Rₘᵢₙ) / (Rₘₐₓ - R)
- Orthogonal Validation: Validate sensor output by comparing with enzymatic assays that measure NADPH and NADP+ in cell extracts, or by using fluorescence lifetime imaging (FLIM) of intrinsic NAD(P)H, which provides independent metabolic information [51].
The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential Reagents and Resources for NADPH/NADP+ Biosensor Research
| Reagent / Resource | Function / Description | Example Use Case | Key Considerations |
|---|---|---|---|
| NAPstar Plasmids [3] | Genetically encoded biosensors for NADP redox state. | Real-time, subcellular monitoring of NADPH/NADP+ dynamics in live cells. | Select variants (e.g., NAPstar3) for pool-size independence and compatibility with FLIM. |
| Peredox-mCherry [3] | Parental scaffold for NADH/NAD+ sensing. | Serves as a control and illustrates engineering progress toward NADP+ specificity. | Critical for understanding the structure-function relationship in sensor engineering. |
| Dual-Sensing Biosensor [53] | Biosensor responsive to both NADPH and a specific metabolite (e.g., L-threonine). | High-throughput screening of microbial strains for metabolic engineering. | Enables growth-coupled selection strategies using FACS. |
| FLIM / trFAIM [51] | Fluorescence Lifetime Imaging Microscopy / time-resolved Fluorescence Anisotropy Imaging. | Label-free metabolic profiling via intrinsic NAD(P)H fluorescence; validates binding configurations. | Provides orthogonal data to confirm findings from genetically encoded biosensors. |
| H₂O₂ & DTT/Rotenone | Chemical oxidant and reductant/inhibitor. | Used for in vivo two-point calibration of biosensor signal. | Concentrations must be optimized for each cell type to achieve full clamping without excessive toxicity. |
Integrated Workflow and Visual Synthesis
The following diagram synthesizes the logical workflow for addressing the key technical challenges in NADPH/NADP+ biosensor applications, from initial characterization to data interpretation.
Workflow for Technical Challenges
The accurate measurement of NADPH/NADP+ redox states using genetically encoded biosensors is pivotal for advancing our understanding of cellular metabolism. By systematically addressing the technical challenges of pool size dependence, pH sensitivity, and in vivo calibration through the protocols and frameworks outlined here, researchers can significantly enhance the reliability and biological relevance of their data. The integration of robust sensor characterization, careful experimental design, and orthogonal validation methods forms the foundation for generating high-quality, interpretable data. This rigorous approach will ultimately accelerate discoveries in redox biology and the development of strategies for the dynamic regulation of NADPH/NADP+ balance in both basic research and therapeutic applications.
The redox balance of the NADPH/NADP+ couple represents a fundamental regulatory node in central metabolism, influencing anabolic capacity, antioxidative defense, and cellular signaling across diverse biological systems [3]. In metabolic engineering, dynamic regulation of this cofactor pair is crucial for optimizing the production of high-value compounds, particularly those requiring substantial reducing power, such as amino acids, biofuels, and nutraceuticals [53]. Traditional static engineering approaches often fail to address the dynamic nature of cellular metabolism, leading to metabolic imbalances, suboptimal productivity, and impaired cellular fitness [54].
The convergence of genetically encoded biosensors and CRISPR interference (CRISPRi) technologies has created powerful platforms for addressing these challenges. Biosensors provide real-time, in vivo monitoring of metabolic states with subcellular resolution, while CRISPRi enables programmable, titratable control of gene expression across the genome [55] [56]. This combination allows for the creation of integrated workflows that can identify genetic bottlenecks and optimize metabolic fluxes at unprecedented scale and resolution, moving beyond static pathway engineering toward dynamic, self-regulating microbial systems [36].
This application note details practical methodologies for implementing biosensor-assisted CRISPRi screening, with particular emphasis on applications relevant to NADPH/NADP+ redox balance. We provide experimental protocols, reagent specifications, and workflow visualizations to facilitate adoption of these techniques in metabolic engineering research and development.
Research Reagent Solutions
Table 1: Essential research reagents for biosensor-CRISPRi workflows
| Reagent Category | Specific Examples | Function and Application |
|---|---|---|
| NAD(P)H Biosensors | NAPstar family [3], iNap sensors [3] | Real-time monitoring of NADPH/NADP+ redox state with subcellular resolution |
| CRISPRi Components | dCas9/dCpf1, sgRNA expression systems [55] [56] | Programmable transcriptional repression of target genes |
| Titratable Repression Systems | Mismatch sgRNA libraries [55] | Enable graded gene knockdown essential for tuning metabolic pathways |
| Metabolite Biosensors | LldR-based lactate sensor [56], GlcN6P biosensors [54] | Product-specific sensing for pathway-specific optimization |
| Fluorescent Reporters | GFP, mCherry, eGFP [55] [36] | Quantitative readout for biosensor activation and sorting |
| Selection Platforms | FACS [53] [56] | High-throughput isolation of high-producing strains |
NADPH/NADP+ Biosensor Characterization and Implementation
Biosensor Selection and Properties
The NAPstar family of genetically encoded biosensors represents a significant advancement for monitoring NADPH/NADP+ redox states across diverse biological systems [3]. These sensors were developed through rational engineering of the bacterial transcriptional repressor Rex and offer several advantages over previous generations, including reduced pH sensitivity, compatibility with fluorescence lifetime imaging (FLIM), and specificity for the NADP(H) pool over NAD(H) [3].
Key characterization data for the NAPstar biosensor family:
Table 2: Performance characteristics of NADP(H) biosensors
| Biosensor Variant | Kd(NADPH) (μM) | Kr(NADPH/NADP+) | Dynamic Range | Key Applications |
|---|---|---|---|---|
| NAPstar1 | 0.9 | 0.006 | ~2.5-fold | Detection of highly oxidized NADP pools |
| NAPstar3 | 2.4 | 0.016 | ~2.5-fold | General purpose applications |
| NAPstar6 | 11.6 | 0.077 | ~2.5-fold | Detection of highly reduced NADP pools |
| iNap Sensors | Various | N/A | ~3-fold | Previous generation with lower brightness |
Biosensor Implementation Protocol
Materials:
- NAPstar plasmid DNA (Addgene)
- Appropriate microbial expression vectors
- Fluorescence spectrometer or flow cytometer
- Confocal microscope with FLIM capability (optional)
Procedure:
- Subcellular Targeting: Clone NAPstar variants into destination vectors containing localization sequences (e.g., mitochondrial, cytosolic) for compartment-specific redox measurements.
- Transformation: Introduce constructs into host strains using appropriate methods (electroporation for bacteria, lithium acetate for yeast).
- Calibration: Perform in vivo calibration using dithiothreitol (DTT) and diamide treatments to establish fully reduced and oxidized ratios, respectively.
- Validation: Confirm sensor functionality via fluorescence excitation ratio imaging (~400 nm/~488 nm) normalized to reference signal (mCherry).
- Application: Monitor NADP redox state changes under experimental conditions (e.g., nutrient shifts, oxidative stress, pathway manipulation).
Biosensor-Assisted CRISPRi Screening Workflow
The integration of biosensors with CRISPRi libraries enables high-throughput identification of genetic targets that optimize NADPH-dependent bioproduction. The following workflow diagram illustrates the key steps in this process:
Diagram 1: Biosensor-assisted CRISPRi screening workflow for strain optimization.
Titratable CRISPRi Library Design and Construction
Mismatch CRISPRi technology enables fine-tuning of gene repression levels, which is essential for optimizing metabolic pathways without complete gene knockout that could impair cell viability [55].
Materials:
- dCas9 expression vector (constitutive or inducible)
- sgRNA scaffold backbone
- Oligonucleotide pools for target gene sgRNAs
- Molecular biology reagents for cloning
Procedure:
- Target Selection: Identify genes of interest in central carbon metabolism, cofactor biosynthesis, and competing pathways.
- sgRNA Design: Design fully complementary sgRNAs plus variants with 1-2 consecutive mismatches in the seed region (positions 5-14 of the spacer) to create a repression gradient [55].
- Library Construction:
- Synthesize oligonucleotide pool encoding all sgRNA variants
- Amplify pool via PCR with appropriate overhangs
- Clone into sgRNA expression vector via Golden Gate assembly or ligation
- Transform into expression host and verify library diversity via sequencing
- Library Validation: Test repression efficiency of individual sgRNA variants against target genes using qRT-PCR or reporter assays.
High-Throughput Screening Protocol
Materials:
- Biosensor-equipped CRISPRi library strains
- Fluorescence-activated cell sorter (FACS)
- Deep well plates or bioreactor systems
- Next-generation sequencing platform
Procedure:
- Culture Preparation: Inoculate library strains in appropriate medium and grow to mid-log phase.
- Induction: If using inducible systems, add inducer to activate dCas9 and sgRNA expression.
- Incubation: Allow sufficient time for gene repression and metabolic adaptation (typically 12-48 hours).
- Fluorescence Detection: Measure biosensor signal via flow cytometry or plate reader.
- Cell Sorting: Isolate populations with desired biosensor response (e.g., high fluorescence for reduced NADP state) using FACS.
- Recovery and Expansion: Culture sorted cells to generate sufficient biomass for downstream analysis.
- Target Identification: Extract genomic DNA from sorted populations, amplify sgRNA regions, and sequence to identify enriched targets.
Table 3: Example screening outcomes from biosensor-CRISPRi applications
| Host Organism | Target Product | Identified Targets | Production Improvement | Citation |
|---|---|---|---|---|
| Zymomonas mobilis | d-lactate | ZMO1323, ZMO1530 | 15-21% increase | [56] |
| Escherichia coli | p-coumaric acid | pfkA, ptsI | 40.6% increase (1308.6 mg/L) | [55] |
| Escherichia coli | butyrate | sucA, ldhA | 19.0-25.2% increase | [55] |
| Pseudomonas putida | isoprenol | Multiple targets from genome-wide screen | 36-fold increase (~900 mg/L) | [36] |
Case Study: Redox Imbalance-Driven L-Threonine Production
The Redox Imbalance Forces Drive (RIFD) strategy exemplifies the power of combining biosensors with targeted metabolic engineering for NADPH-dependent products [53]. This approach deliberately creates NADPH excess through "open source and reduce expenditure" strategies, then harnesses this imbalance to drive metabolic flux toward target compounds.
Diagram 2: Redox imbalance forces drive (RIFD) strategy for L-threonine production.
RIFD Implementation Protocol
Materials:
- L-threonine production strain (e.g., E. coli TN)
- MAGE oligonucleotides for targeted evolution
- NADPH and L-threonine dual-sensing biosensor
- FACS instrumentation
Procedure:
- NADPH Enhancement: Implement three "open source" strategies:
- Express cofactor-converting enzymes (e.g., NADH kinase)
- Introduce heterologous NADPH-dependent enzymes
- Enhance native NADPH synthesis pathways (e.g., pentose phosphate pathway)
- Consumption Reduction: Knock down non-essential NADPH-consuming reactions.
- Evolution: Use multiplex automated genome engineering (MAGE) to introduce combinatorial mutations in redox-related genes.
- Screening: Employ dual-sensing biosensor to simultaneously monitor NADPH and L-threonine levels.
- Isolation: Use FACS to isolate strains with both high NADPH and high L-threonine production.
- Validation: Confirm L-threonine production yields via HPLC in bench-scale bioreactors.
Results: This approach yielded a high-producing strain with titer of 117.65 g/L L-threonine and yield of 0.65 g/g glucose [53].
Troubleshooting and Technical Considerations
Biosensor Limitations and Optimization
- Dynamic Range: NAPstar sensors offer ~2.5-fold dynamic range; ensure measurement systems are sufficiently sensitive [3].
- Expression Effects: High biosensor expression may buffer metabolite pools; use moderate expression levels and include empty vector controls.
- Cross-reactivity: Validate NADPH specificity in your host system, as some sensors show affinity for NADH at high concentrations [3].
CRISPRi Library Quality Control
- Coverage: Maintain >1000x library coverage throughout screening to prevent bottleneck effects.
- Repression Efficiency: Validate knockdown efficiency for a subset of targets before full screening.
- Off-target Effects: Include control sgRNAs and validate findings with complementary approaches (e.g., gene deletions).
Screening Optimization
- Sorting Gates: Use appropriate gating strategies based on biosensor dynamics; avoid overly stringent gates that may reduce diversity.
- Culture Conditions: Maintain environmental control throughout screening to prevent non-genetic adaptations.
- Replicate Screening: Perform independent biological replicates to identify consistently enriched targets.
The integration of genetically encoded biosensors with CRISPRi library technologies creates a powerful platform for optimizing NADPH/NADP+ balance and enhancing microbial production of valuable compounds. The workflows described herein enable researchers to move beyond static metabolic engineering toward dynamic, self-regulating systems that maintain redox balance while maximizing product yield. As biosensor specificity and CRISPRi precision continue to improve, these integrated approaches will play an increasingly important role in accelerating the development of robust microbial cell factories for industrial biotechnology.
Benchmarking Performance: A Comparative Analysis of NADP(H) Biosensor Platforms
The NADPH/NADP+ redox couple constitutes a fundamental component of cellular redox metabolism, serving as a crucial electron donor in anabolic pathways and antioxidative defense systems across all living organisms [3]. Research into the dynamic regulation of NADPH/NADP+ balance has been revolutionized by the development of genetically encoded biosensors, which enable real-time monitoring of subcellular redox states with unprecedented resolution [3]. The recent introduction of the NAPstars biosensor family addresses long-standing limitations in the field, offering researchers powerful new tools to investigate redox metabolism across yeast, plants, and mammalian cell models [3]. This application note provides a comprehensive technical comparison of these advanced biosensors, detailing their key performance metrics and experimental protocols for researchers and drug development professionals working at the intersection of redox biology and metabolic engineering.
Table 1: Core Components of a NADPH/NADP+ Biosensing System
| Component | Description | Function in Biosensing |
|---|---|---|
| Bioreceptor | Rex NADH/NAD+-binding domain (engineered) | Biological recognition element that specifically binds NADPH/NADP+ |
| Transducer | Circularly permuted T-Sapphire (cpTS) | Converts binding event into measurable fluorescent signal |
| Reference Element | mCherry fluorescent protein | Provides ratiometric internal control for quantification |
| Expression System | Genetically encoded plasmid | Enables targeted subcellular localization and expression in model organisms |
Performance Metrics Comparison of NADPH/NADP+ Biosensors
The quantitative characterization of biosensor performance is essential for experimental design and data interpretation. The following section provides a detailed comparison of key parameters for the NAPstars biosensor family.
Table 2: Key Performance Metrics of NAPstars Biosensor Variants
| Biosensor Variant | Dynamic Range (Ratio Change) | Kd(NADPH) (μM) | Kr(NADPH/NADP+) | Specificity (NADPH vs. NADH) | Response Time to Equilibrium |
|---|---|---|---|---|---|
| NAPstar1 | ~2.5-fold | 0.9 | ~0.006 | ~27x higher affinity for NADPH | Not explicitly stated |
| NAPstar2 | ~2.5-fold | Data not specified | Data not specified | Data not specified | Not explicitly stated |
| NAPstar3 | ~2.5-fold | Data not specified | Data not specified | ~27x higher affinity for NADPH | Not explicitly stated |
| NAPstar6 | ~2.5-fold | 11.6 | ~0.077 | ~21x higher affinity for NADPH | Not explicitly stated |
| NAPstar7 | ~2.5-fold | Data not specified | Data not specified | ~10x higher affinity for NADPH | Not explicitly stated |
| jAspSnFR3 (Aspartate Sensor) | 20-fold | 50 (for aspartate) | Not applicable | >100x preference for aspartate over glutamate | 130 ms |
Analysis of Performance Metrics
The NAPstars family exhibits a consistent dynamic range of approximately 2.5-fold increase in fluorescence upon NADPH binding, which is comparable to their predecessor Peredox [3]. This range is sufficient for detecting physiological changes in NADPH/NADP+ ratios, though it is notably lower than the 20-fold dynamic range demonstrated by the jAspSnFR3 aspartate sensor [57]. Affinity varies significantly across variants, with NAPstar1 showing the highest affinity (Kd(NADPH) = 0.9 μM) and NAPstar6 the lowest (Kd(NADPH) = 11.6 μM) [3]. This affinity range allows researchers to select sensors appropriate for expected cellular NADPH concentrations, with higher affinity sensors better suited for detecting low abundance pools.
Specificity profiling reveals that all NAPstars show substantially higher affinity for NADPH over NADH (by factors of 10 to 27), a crucial feature for accurate NADPH-specific monitoring without cross-talk from the abundant NADH pool [3]. The sensors demonstrate minimal response to NAD+, confirming their specificity for the phosphorylated redox couple [3]. While explicit response times were not provided for NAPstars, their design based on Peredox suggests relatively rapid kinetics suitable for monitoring metabolic oscillations, such as those observed during the yeast cell cycle [3].
Experimental Protocols for Biosensor Implementation
Protocol 1: In Vitro Characterization of NAPstars Biosensors
Purpose: To determine the affinity, specificity, and dynamic range of NAPstars biosensors using purified recombinant protein.
Materials:
- Recombinant NAPstar protein (expressed in E. coli and purified)
- NADPH, NADP+, NADH, NAD+ standards (prepare fresh solutions in appropriate buffer)
- Assay buffer (e.g., 50 mM Tris-HCl, 100 mM NaCl, pH 7.4)
- Fluorescence spectrophotometer with excitation and emission scanning capabilities
- Cuvettes or microplate reader compatible with ratiometric measurements
Procedure:
- Express and purify NAPstar proteins with His-tags using standard nickel affinity chromatography.
- Prepare a master solution of NAPstar protein diluted in assay buffer to a consistent concentration.
- For affinity determination, titrate NADPH (0.01-1000 μM) in the presence of constant NADP+ (150 μM).
- Measure fluorescence excitation spectra (400 nm excitation, 515 nm emission for TS) and emission (580 nm for mCherry) after each addition.
- Calculate the TS/mCherry ratio for each NADPH concentration and fit the data to a binding isotherm to determine Kd(NADPH).
- Repeat titrations with NADH (in presence of NAD+) and NADP+ to determine specificity.
- For redox state measurements, maintain a constant total NADP pool (100, 300, or 500 μM) while varying the NADPH/NADP+ ratio from 0.001 to 5.
Validation: NAPstar sensors should maintain consistent responses across different total NADP pool sizes, confirming their reporting of authentic NADPH/NADP+ ratios rather than absolute concentrations [3].
Protocol 2: Monitoring NADP Redox State in Live Mammalian Cells
Purpose: To implement NAPstars for real-time monitoring of subcellular NADP redox dynamics in response to oxidative challenge.
Materials:
- Mammalian cell line (e.g., HEK293, HeLa)
- NAPstar expression plasmid (subcellular targeting sequences can be added)
- Transfection reagent (e.g., lipofectamine)
- Live-cell imaging medium (without phenol red)
- Confocal or epifluorescence microscope with environmental control (37°C, 5% CO₂)
- Image analysis software (e.g., ImageJ, MATLAB)
- Oxidative stress inducers (e.g., H₂O₂) and antioxidants (e.g., N-acetylcysteine)
Procedure:
- Culture cells on glass-bottom dishes or plates suitable for microscopy.
- Transfect cells with NAPstar plasmid using standard transfection protocols.
- 24-48 hours post-transfection, replace medium with live-cell imaging medium.
- Image cells using appropriate filter sets: 400/515 nm for TS and 580/630 nm for mCherry.
- Acquire baseline ratiometric images (TS/mCherry) for 5-10 minutes.
- Apply oxidative challenge (e.g., 100-500 μM H₂O₂) or other metabolic perturbations directly to cells while continuing time-lapse imaging.
- Analyze ratio images after background subtraction, normalizing to baseline values.
- For compartment-specific analysis, use segmentation tools to isolate signals from cytosol, mitochondria, or other target organelles.
Troubleshooting: Ensure proper sensor expression levels, as excessive expression may buffer endogenous metabolites and cause artifacts. Include control experiments with NADP+-insensitive mutant (NAPstarC) to confirm specificity of observed changes [3].
Signaling Pathways and Experimental Workflows
The following diagrams illustrate the core NADP redox coupling and experimental workflows for biosensor characterization and application.
NADP Redox Coupling in Cellular Metabolism
Diagram 1: NADP Redox Coupling. Metabolic pathways including pentose phosphate pathway (PPP), fatty acid oxidation (FAO), malic enzyme (ME1), and isocitrate dehydrogenase (IDH) generate NADPH. NADPH is consumed by glutathione (GSH) and thioredoxin (TXN) systems for reactive oxygen species (ROS) detoxification, regenerating NADP+.
Biosensor Characterization and Application Workflow
Diagram 2: Biosensor Workflow. The comprehensive pipeline for biosensor development and application begins with rational design, proceeds through in vitro characterization, cellular implementation, live-cell imaging, and culminates in data analysis for biological insights.
Research Reagent Solutions
The following table details essential materials and reagents for implementing NADPH/NADP+ biosensing studies.
Table 3: Essential Research Reagents for NADPH/NADP+ Biosensing Studies
| Reagent/Category | Specific Examples | Function/Application |
|---|---|---|
| Biosensor Plasmids | NAPstars (1, 2, 3, 6, 7), NAPstarC (control), jAspSnFR3 | Core sensing elements for NADPH/NADP+ or related metabolites |
| Expression Systems | Bacterial (E. coli), Yeast, Plant, Mammalian cell lines | Host organisms for biosensor expression and validation |
| Microscopy Systems | Confocal microscopes, Epifluorescence systems, FLIM-capable instruments | Detection platforms for ratiometric or lifetime imaging |
| Electrode Platforms | Screen-printed electrodes (SPEs), Thin-film electrodes | Electrochemical sensing platforms for complementary approaches |
| Metabolic Modulators | H₂O₂, N-acetylcysteine, hypoxia chambers, mitochondrial inhibitors | Tools for perturbing redox state to validate sensor response |
| Validation Tools | LC-MS/MS systems, Fluorescence-activated cell sorting (FACS) | Orthogonal methods to confirm biosensor accuracy |
The development and characterization of the NAPstars biosensor family represents a significant advancement in our ability to monitor NADPH/NADP+ redox dynamics with high specificity and subcellular resolution. The quantitative performance metrics presented in this application note provide researchers with critical information for selecting appropriate sensor variants for specific experimental contexts. The detailed protocols enable straightforward implementation across various model systems, from in vitro biochemical characterization to live-cell imaging applications.
Future directions in biosensor development will likely focus on expanding the color palette for multiplexed imaging, improving dynamic range further, and engineering sensors with modified affinities for specific subcellular compartments. The integration of these biosensors with other metabolic probes and orthogonal validation methods will provide increasingly comprehensive views of cellular redox regulation. For drug development professionals, these tools offer unprecedented capability to screen compounds for effects on redox metabolism, potentially identifying novel therapeutics for conditions involving redox imbalance, including metabolic diseases, cancer, and age-related disorders.
The NADPH/NADP+ redox couple constitutes a central metabolic node, differentially regulating redox signalling, antioxidative defence, and anabolic processes across subcellular compartments. Understanding the dynamic regulation of this balance requires tools that can capture these changes in real-time within living systems. Genetically encoded fluorescent biosensors have revolutionized this field by enabling specific, non-invasive monitoring of metabolic states with spatiotemporal resolution unattainable by traditional biochemical methods [30]. This guide provides a structured framework for selecting and implementing the most appropriate NADP redox biosensors for your specific research objectives, focusing on the latest technological advances and their practical applications in drug development and basic research.
Biosensor Selection Framework
Understanding Biosensor Operating Principles
Genetically encoded biosensors translate biochemical events into measurable fluorescence signals through several distinct mechanisms:
- Ratiometric Excitation Sensors: These sensors, including the NAPstar family, exhibit shifts in excitation peaks upon analyte binding, allowing quantification through excitation ratioing, which minimizes artifacts from variable expression levels or sample thickness [3] [58].
- FRET-Based Sensors: Employ two fluorescent proteins where analyte-induced conformational changes alter Förster Resonance Energy Transfer efficiency, detectable as emission ratio changes [59] [58].
- Single-Fluorophore Intensity Sensors: Utilize circularly permuted fluorescent proteins whose brightness changes with analyte binding, offering simpler implementation but greater susceptibility to environmental artifacts [58] [30].
- FLINC-Based Sensors: Leverage binding-induced changes in fluorescence fluctuations, quantifiable through stochastic optical fluctuation imaging to achieve superresolution imaging of biochemical activities [60].
Quantitative Comparison of NADP Redox Biosensors
Table 1: Characteristics of Genetically Encoded NADP Redox Biosensors
| Biosensor Name | Sensing Target | Dynamic Range | Key Advantages | Reported Applications |
|---|---|---|---|---|
| NAPstar family [3] | NADPH/NADP+ ratio | ~5000-fold (ratios 0.001-5) | Broad dynamic range, subcellular resolution, pH stability, compatible with FLIM | Yeast, plants, mammalian cells; oxidative stress studies |
| iNap [30] | NADPH concentration | Not specified | Specific NADPH detection | Developmental biology, redox homeostasis |
| Apollo-NADP+ [30] | NADP+ concentration | Not specified | Specific NADP+ detection | Subcellular NADP+ dynamics |
| NADP-Snifit [30] | NADP(H) levels | Not specified | FRET-based semisynthetic design | Live-cell NADP(H) monitoring |
| Peredox [3] | NADH/NAD+ ratio | ~2.5 (spectroscopic) | Parent scaffold for NAPstars | NAD redox state monitoring |
| SoNar [30] | NAD+/NADH ratio | Not specified | High sensitivity to NADH/NAD+ | Cellular energy metabolism |
Table 2: Performance Characteristics of NAPstar Variants
| NAPstar Variant | Kr(NADPH/NADP+) | Specificity vs NADH | pH Sensitivity | Recommended Use Cases |
|---|---|---|---|---|
| NAPstar1 | 0.9 µM | 27x higher affinity for NADPH | Low | High-resolution detection in oxidized compartments |
| NAPstar3 | 2.1 µM | 24x higher affinity for NADPH | Low | General purpose applications |
| NAPstar6 | 11.6 µM | 21x higher affinity for NADPH | Low | Detection in reduced environments |
| NAPstar7 | 5.7 µM | 12x higher affinity for NADPH | Low | Intermediate redox state monitoring |
| NAPstarC | No binding | No nucleotide binding | Low | Negative control experiments |
Experimental Protocols
Protocol 1: Implementing NAPstars for Subcellular NADP Redox Monitoring
Principle: NAPstars are genetically encoded biosensors derived from Peredox-mCherry, with engineered Rex domains that specifically favor NADP over NAD binding. They function as ratiometric excitation sensors, with TS (T-Sapphire) fluorescence increasing with NADPH binding, normalized against mCherry reference signal [3].
Workflow:
- Sensor Selection and Expression: Choose appropriate NAPstar variant based on expected NADP redox state (see Table 2). Express in target cells using suitable vectors (e.g., lentiviral for mammalian cells, Agrobacterium for plants).
- Calibration: Perform in vitro calibration with recombinant protein using 0-100 µM NADPH in presence of constant 150 µM NADP+ to establish standard curve.
- Live-Cell Imaging: Image using dual-excitation fluorescence microscopy (TS excitation: ~400 nm; mCherry excitation: ~570 nm) with emission at 515 nm and 610 nm respectively.
- Image Analysis: Calculate ratio of TS/mCherry fluorescence intensities pixel-by-pixel. Convert to NADPH/NADP+ ratio using calibration curve.
- Validation: Confirm specificity using NADP+ titrations and test with NADH/NAD+ to verify minimal cross-reactivity.
Protocol 2: Multiplexed Biosensing of Metabolic Networks
Principle: Simultaneous monitoring of NADP redox state with related metabolic parameters using spectrally distinct biosensors [58].
Workflow:
- Sensor Pair Selection: Choose biosensors with minimal spectral overlap (e.g., NAPstars with green/red sensors for calcium or cAMP).
- Expression Optimization: Titrate DNA ratios to achieve balanced expression of all sensors.
- Spectral Unmixing: Acquire images with appropriate filter sets and apply linear unmixing algorithms to resolve individual signals.
- Temporal Correlation: Analyze dynamics across different metabolic parameters to identify regulatory relationships.
- Pathway Modulation: Apply specific inhibitors (e.g., glutathione system inhibitors) to dissect contribution of antioxidant pathways [3].
Research Reagent Solutions
Table 3: Essential Reagents for NADP Redox Biosensing
| Reagent Category | Specific Examples | Function/Purpose | Implementation Notes |
|---|---|---|---|
| Biosensor Plasmids | NAPstar1-7, iNap, SoNar | Enable specific NADP redox state monitoring | Select variant based on dynamic range requirements; NAPstars recommended for ratio measurements [3] |
| Expression Systems | Lentiviral vectors, Cell-specific promoters | Controlled biosensor delivery | Use constitutive promoters for steady-state studies; inducible systems for developmental tracking |
| Calibration Reagents | NADPH, NADP+, NADH, NAD+ | In vitro sensor characterization and standard curves | Essential for quantitative interpretation; include in all experimental setups [3] |
| Pathway Modulators | BSO (glutathione synthesis inhibitor), Auranofin (thioredoxin reductase inhibitor) | Dissect antioxidant pathway contributions | Revealed glutathione system as primary mediator of antioxidative electron flux [3] |
| Imaging Tools | Fluorescence microscopes with ratiometric capability, FLIM systems | Biosensor signal detection | FLIM compatible with NAPstars for alternative readout modality [3] |
Advanced Applications and Integration
Integration with Superresolution Techniques
The combination of biosensors with advanced imaging modalities enables unprecedented resolution of metabolic compartmentalization:
- FLINC-Based Biosensors: Utilize fluorescence fluctuation increases by contact between Dronpa and TagRFP-T to achieve superresolution mapping of biochemical activities, resolving features as small as 116 nm [60].
- FLIM Compatibility: NAPstars support fluorescence lifetime imaging microscopy (FLIM), providing an alternative readout modality that is less susceptible to concentration effects and photobleaching [3].
- pcSOFI Imaging: Photochromic Stochastic Optical Fluctuation Imaging quantifies fluorescence fluctuation strength at sub-pixel resolution, enabling superresolution activity mapping [60].
Chemogenetic FRET Pairs for Enhanced Dynamic Range
Recent advances in chemogenetic biosensor design offer significantly improved performance characteristics:
- ChemoX Platform: Engineered FRET pairs with near-quantitative efficiency (≥94%) created through reversible interaction between fluorescent proteins and fluorescently labeled HaloTag [59].
- Spectral Tunability: Acceptors can be tuned using different rhodamine fluorophores (JF525 to JF669) while maintaining high FRET efficiency [59].
- Implementation: Particularly valuable for multiplexed imaging and when extremely large dynamic ranges are required for detecting subtle metabolic changes.
The strategic selection of biosensors tailored to specific experimental goals is paramount for advancing our understanding of NADPH/NADP+ dynamics in living systems. The development of sophisticated tools like the NAPstar family with expanded dynamic ranges and specificity, combined with advanced imaging modalities and multiplexing approaches, provides unprecedented opportunities to decode the complex regulation of redox metabolism in health and disease. By following the structured framework presented in this guide, researchers can effectively match biosensor capabilities to their specific research questions, accelerating discovery in metabolic research and drug development.
The dynamic regulation of the NADPH/NADP+ balance is a central metabolic process, critical for maintaining redox homeostasis, supporting biosynthetic reactions, and enabling cellular antioxidant defense [3] [30]. Genetically encoded biosensors have revolutionized our ability to monitor these dinucleotide pools in living cells with high spatiotemporal resolution. However, the fidelity of these tools must be rigorously established through correlation with established gold-standard techniques. This Application Note provides detailed protocols for validating NADPH/NADP+ biosensor readouts by correlating them with Fluorescence Lifetime Imaging Microscopy (FLIM), High-Performance Liquid Chromatography (HPLC), and key physiological phenotypes, providing a robust framework for confident data interpretation in pharmacological and metabolic research.
The Biosensor Toolbox for NADP(H) Monitoring
A suite of genetically encoded biosensors has been developed to probe the NADP(H) pool, each with distinct properties, affinities, and applications. Key performance characteristics of contemporary NADP(H) biosensors are summarized in Table 1.
Table 1: Key Characteristics of Genetically Encoded NADP(H) Biosensors
| Sensor Name | Target | Dynamic Range / Kd / C50 | Key Features and Validation |
|---|---|---|---|
| NAPstars [3] | NADPH/NADP+ Ratio | Kratio(NADPH/NADP+): 0.001 to 5 | Rational design from Peredox; compatible with FLIM; pH-resistant; validated in yeast, plants, mammalian cells. |
| iNap [9] | NADPH | Kd: ~1.3 µM to ~29 µM (iNap1-4) | Ratiometric; pH-resistant; wide dynamic range; used with flow cytometry; subcellular targeting. |
| NERNST [32] | NADP(H) Redox Potential (ENADP(H)) | N/A | Ratiometric; based on roGFP2 fused to NTRC; specific to NADP(H) redox states. |
| mBFP [61] | NADPH | Kd: 0.64 mM | Oxygen-independent; catalytic activity (SDR); used in bacteria (e.g., C. glutamicum, E. coli). |
| NADP-Snifit [31] | NADPH/NADP+ Ratio | r50 (Ratio): 30 | Semisynthetic FRET sensor; pH-insensitive; long-wavelength excitation; requires dye labeling. |
The selection of an appropriate biosensor depends on the specific research question. For instance, the NAPstar family is ideal for detecting subtle redox changes across a wide range and is uniquely compatible with FLIM [3], while iNap sensors offer a range of affinities suitable for different subcellular compartments [9]. NERNST directly reports the thermodynamic redox potential, ENADP(H), providing a direct measure of cellular energy status [32].
Correlative Experimental Protocols
Protocol: Correlation with Fluorescence Lifetime Imaging (FLIM)
FLIM provides a robust, concentration-independent readout of fluorescence decay kinetics, which can be influenced by biosensor conformational changes upon metabolite binding.
- Principle: The fluorescence lifetime of a fluorophore is sensitive to its molecular environment. A biosensor's shift between NADPH-bound and free states can alter the lifetime, enabling a direct measurement of the redox state without ratiometric imaging [3].
- Workflow: The following diagram illustrates the key steps for performing a FLIM-based validation of biosensor readouts.
- Detailed Methodology:
- Cell Preparation and Imaging:
- Seed cells (e.g., HeLa, yeast) expressing the biosensor (e.g., NAPstar) in glass-bottom dishes.
- Acquire time-lapse ratiometric images (e.g., 400nm/500nm ex for TS, emission at >515nm for NAPstars) on a confocal microscope to establish a baseline ratio (R) [3].
- Immediately transfer the dish to a FLIM-equipped multiphoton or confocal microscope. Use a pulsed laser (e.g., Ti:Sapphire) tuned to the excitation wavelength (e.g., 820 nm for two-photon excitation of cpFP-based sensors). Collect emitted photons using a high-sensitivity detector (e.g., hybrid PMT) and a time-correlated single-photon counting (TCSPC) module.
- Lifetime Analysis:
- Fit the fluorescence decay curve for each pixel to a multi-exponential model (e.g., ( I(t) = α₁exp(-t/τ₁) + α₂exp(-t/τ₂) + C )) using specialized software (e.g., SPCImage, SymPhoTime).
- Calculate the amplitude-weighted average lifetime ( τ_{avg} = (α₁τ₁ + α₂τ₂)/(α₁ + α₂) ) to generate a lifetime map.
- Correlation and Validation:
- For the same field of view and time point, plot the pixel-by-pixel average fluorescence lifetime (τ) against the ratiometric value (R).
- A strong, significant correlation (e.g., Pearson's r > |0.7|) validates that the ratiometric shift reports a genuine molecular event (conformational change) related to the NADP(H) state, as demonstrated with the NAPstar biosensor [3].
- Cell Preparation and Imaging:
Protocol: Correlation with HPLC
HPLC provides absolute, quantitative data on NADPH and NADP+ concentrations, serving as a biochemical ground truth.
- Principle: Separates and quantifies NADPH and NADP+ from cell extracts based on retention time, using UV/Vis or mass spectrometry detection.
- Workflow: The following diagram outlines the parallel processing of samples for biosensor imaging and HPLC analysis.
- Detailed Methodology:
- Parallel Sample Preparation:
- Grow and treat cells in multiple identical culture plates or flasks.
- For one set, perform live-cell ratiometric imaging of the biosensor and record the average ratio from a population of cells.
- For the parallel set, rapidly aspirate the medium and quench metabolism immediately. For intracellular NADP(H), this is critical due to fast turnover. Use a dedicated quenching solution (e.g., cold acidic acetonitrile/methanol, liquid N₂) to freeze metabolic activity [61].
- Metabolite Extraction:
- Scrape quenched cells in the extraction solvent.
- Perform multiple cycles of freeze-thaw or sonication on ice to ensure complete metabolite release.
- Centrifuge at high speed (e.g., 16,000 × g, 10 min, 4°C) to pellet protein and cell debris. Transfer the clear supernatant to a new tube and dry it using a speed vacuum concentrator. Resuspend the pellet in the HPLC mobile phase.
- HPLC Analysis:
- Use a reverse-phase C18 column or a HILIC column for separation.
- Employ a gradient elution with a mobile phase such as potassium phosphate buffer (e.g., 100 mM, pH 6.0) and methanol.
- Detect NADPH and NADP+ by their absorbance at 340 nm. Quantify concentrations by comparing peak areas to a standard curve of pure NADPH and NADP+ run on the same day.
- Data Correlation:
- Plot the biosensor ratiometric value (or FRET ratio) against the HPLC-measured [NADPH]/[NADP+] ratio or the absolute concentration of NADPH. A linear relationship across a range of metabolic perturbations (e.g., glucose addition, paraquat treatment) confirms the quantitative accuracy of the biosensor, as performed for mBFP in C. glutamicum [61].
- Parallel Sample Preparation:
Protocol: Correlation with Physiological Phenotypes
The ultimate validation of a biosensor's biological relevance is its ability to predict or explain a cellular phenotype.
- Principle: Biosensor readouts are correlated with measurable phenotypic outcomes, such as oxidative stress sensitivity, cell proliferation, or specific metabolic fluxes.
- Workflow: This workflow demonstrates how biosensor measurements can be integrated with phenotypic assays to establish functional relevance.
- Detailed Methodology:
- Perturbation and Concurrent Monitoring:
- Oxidative Stress: Treat cells expressing the NADPH biosensor (e.g., iNap, NERNST) with paraquat or H₂O₂ while monitoring the NADPH level or ENADP(H) in real-time [61] [32]. In parallel, measure reactive oxygen species (ROS) using a fluorescent dye like CM-H₂DCFDA and assess cell viability via propidium iodide staining or a colony-forming assay.
- Metabolic Challenges: Shift cells from a carbon source that generates low NADPH to one that fuels the pentose phosphate pathway (e.g., glucose). Monitor the ensuing increase in biosensor signal (e.g., with iNap [9] or mBFP [61]) and correlate it with the rate of cell proliferation or the secretion of an NADPH-dependent product (e.g., fatty acids, lysine in production strains [61]).
- Genetic Interventions: Knock down or overexpress genes central to NADPH metabolism (e.g., Glucose-6-phosphate dehydrogenase, G6PD; NAD+ kinase, NADK) [9]. Use the biosensor to confirm the expected changes in the NADP(H) pool and correlate these changes with alterations in the global metabolic network, potentially using transcriptomics or proteomics.
- Data Interpretation:
- A successful correlation is demonstrated when the biosensor dynamics (e.g., the rate of NADPH depletion, the steady-state NADPH level) consistently align with the phenotypic severity (e.g., cells with a slower NADPH recovery post-stress show higher mortality). For example, NAPstar sensors revealed the glutathione system as the primary mediator of antioxidative electron flux across eukaryotes, directly linking sensor readouts to a conserved physiological mechanism [3].
- Perturbation and Concurrent Monitoring:
Research Reagent Solutions
Table 2: Essential Reagents and Tools for NADP(H) Biosensor Validation
| Category / Item | Function / Application | Example Use Case |
|---|---|---|
| Biosensor Plasmids | Tool for live-cell imaging of NADP(H) dynamics. | NAPstars [3], iNap [9], NERNST [32] |
| FLIM System | Microscope for fluorescence lifetime measurement. | Validates biosensor mechanism; provides concentration-independent readout [3]. |
| HPLC with UV/Vis Detector | Instrument for absolute quantification of metabolites. | Measures absolute NADPH/NADP+ concentrations from cell extracts for calibration [61]. |
| Chemical Reductants / Oxidants | Tools to perturb cellular redox state. | DTT (reductant), H₂O₂ (oxidant) to define sensor dynamic range in vivo [32]. |
| Metabolic Modulators | Pharmaceuticals to specifically manipulate pathways. | Paraquat (induces NADPH consumption), G6PD inhibitor (6-AN) to test sensor specificity [9] [61]. |
| Cell Viability Assays | Kits/dyes to measure phenotypic outcomes. | Propidium iodide, MTT, CFU assays to correlate NADPH levels with survival [32]. |
Robust validation is the cornerstone of reliable research using genetically encoded biosensors. The multi-faceted approach outlined here—correlating dynamic biosensor data with FLIM, absolute HPLC quantification, and relevant physiological phenotypes—creates a powerful feedback loop that both verifies the tool's performance and generates profound biological insights. By implementing these protocols, researchers in drug development and metabolic engineering can confidently use NADP(H) biosensors to dissect the dynamic regulation of redox metabolism, identify novel drug targets, and screen for compounds that modulate this critical cellular balance.
The nicotinamide adenine dinucleotide phosphate (NADPH/NADP+) redox couple constitutes a central node in metabolic networks, serving as a crucial electron donor for reductive biosynthesis and antioxidant defense systems [3] [8]. Maintaining the optimal NADPH/NADP+ balance is critical for cellular viability, as an imbalance can disrupt cell growth and the production of high-value chemicals [8]. Historically, a detailed understanding of subcellular NADP redox dynamics was limited by the lack of tools capable of specific, real-time monitoring within living cells [3]. The emergence of genetically encoded biosensors has revolutionized this field, enabling researchers to observe these dynamics with high spatiotemporal resolution [30]. These biosensors are engineered proteins that typically combine a sensing domain, which undergoes a conformational change upon binding the target metabolite, with a fluorescent protein reporter, the emission properties of which change accordingly [30]. This technology was pivotal for the unexpected discovery that the glutathione system, rather than the thioredoxin system, plays the predominant role in mediating antioxidative electron flux across diverse eukaryotes [3].
The NAPstars Biosensor Family: Design and Characterization
Sensor Engineering and Validation
The NAPstar family of biosensors was developed to address significant limitations of previous NADPH sensors, such as pH sensitivity, lack of responsiveness to NADP+, and low signal-to-noise ratio [3]. Researchers used the Peredox-mCherry sensor, originally designed for reporting NADH/NAD+ ratios, as a structural chassis [3]. A rational design strategy was employed, introducing specific mutations known to switch the cofactor specificity of the bacterial transcriptional repressor Rex domain from favoring NADH to favoring NADPH [3]. This resulted in a family of constructs, the NAPstars, which feature a circularly permuted T-Sapphire fluorescent protein nested between two engineered Rex domains, with a C-terminally fused mCherry for signal normalization [3].
Table 1: Key Characteristics of NAPstar Biosensor Variants
| Sensor Variant | Kd(NADPH) (µM) | Kr(NADPH/NADP+) | Dynamic Range | Key Application |
|---|---|---|---|---|
| NAPstar1 | 0.9 | - | ~2.5 | Highly reduced conditions |
| NAPstar3 | - | - | ~2.5 | General cytosolic sensing |
| NAPstar6 | 11.6 | - | ~2.5 | Oxidized conditions |
| iNap | - | - | - | NADPH concentration [30] |
| SoNar | - | - | - | NADH/NAD+ ratio [30] |
In vitro characterization confirmed that NAPstars are ratiometric biosensors responsive to the NADPH/NADP+ ratio, not merely the NADPH concentration [3]. They exhibit a broad dynamic range, covering NADPH/NADP+ ratios from approximately 0.001 to 5, and possess substantially higher affinity for NADPH than for NADH, ensuring specificity for the NADP pool [3]. Furthermore, their design incorporates a Rex dimer within a single polypeptide, making their performance less dependent on sensor concentration compared to other designs [3].
Experimental Protocol: In Vitro Biosensor Characterization
Objective: To purify and characterize the response of a NAPstar biosensor to different NADPH/NADP+ ratios. Materials:
- Recombinant NAPstar protein (e.g., NAPstar1, 2, 3, 6, or 7) purified from E. coli [3]
- NADPH and NADP+ stock solutions
- Suitable buffer (e.g., PBS, pH 7.4)
- Fluorometer or plate reader capable of dual-excitation/emission measurements
Procedure:
- Protein Purification: Express recombinant NAPstar protein in E. coli and purify using affinity chromatography suitable for the chosen fusion tag [3].
- Spectrofluorometric Titration:
- Prepare a solution containing a fixed concentration of NAPstar protein (e.g., 1 µM) and a constant total NADP pool (e.g., 150 µM NADP+).
- Titrate by adding increasing concentrations of NADPH (e.g., 0.01–1000 µM) to create a range of NADPH/NADP+ ratios.
- For each titration point, measure the fluorescence excitation spectrum (400 nm for T-Sapphire) and emission (515 nm for T-Sapphire, ~610 nm for mCherry).
- Calculate the ratio of T-Sapphire/mCherry fluorescence for each condition.
- Data Analysis: Plot the ratiometric signal (T-Sapphire/mCherry) against the NADPH/NADP+ ratio. Fit the data to a binding curve to determine the apparent dissociation constant (Kd) and the dynamic range of the sensor.
Application: Revealing Glutathione's Primary Role in Antioxidant Defense
Experimental Workflow for Pathway Interrogation
The critical discovery of the glutathione system's primacy was made by applying NAPstar biosensors in living cells under oxidative stress while selectively impairing specific antioxidant pathways.
Detailed Protocol: Interrogating Antioxidant Pathways with NAPstars
Objective: To determine the relative contribution of the glutathione and thioredoxin systems to maintaining the NADP redox state during acute oxidative challenge. Materials:
- Cell line (e.g., yeast, mammalian cells) expressing NAPstar biosensor in the cytosol
- Confocal microscope or fluorometer with environmental control
- Auranofin (AF): Selective inhibitor of thioredoxin reductase (TrxR) [3] [62]
- 2,4-Dinitrochlorobenzene (DNCB): Agent that depletes glutathione (GSH) [3] [62]
- Oxidizing agent (e.g., H₂O₂)
- Appropriate cell culture media and reagents
Procedure:
- Cell Preparation: Seed cells stably expressing the cytosopically targeted NAPstar biosensor into appropriate imaging dishes or plates and allow them to adhere and grow to the desired confluency [3].
- Baseline Measurement: Place the sample on the microscope stage. For ratiometric imaging, excite T-Sapphire at ~400 nm and mCherry at ~570 nm, and collect emissions at ~515 nm and ~610 nm, respectively. Acquire images for several minutes to establish a stable baseline NADP redox state, reported as the T-Sapphire/mCherry ratio [3].
- Pathway Inhibition: Treat cells with either:
- Auranofin (e.g., 1–10 µM) to inhibit the thioredoxin system.
- DNCB (e.g., 50–200 µM) to deplete the glutathione pool.
- A combination of both inhibitors.
- Include a vehicle control.
- Monitor the NADP redox state for 30-60 minutes post-inhibition.
- Oxidative Challenge: Expose all treatment groups to a bolus of H₂O₂ (e.g., 100-500 µM) [3]. Continue real-time ratiometric imaging to track the dynamics of the NADPH/NADP+ ratio as the cell responds to the stress.
- Data Analysis: Quantify the fluorescence ratios over time. Key metrics include:
- The magnitude and rate of NADP oxidation immediately after H₂O₂ addition.
- The recovery rate of the reduced NADPH pool.
- The steady-state NADP redox state under each condition.
Key Findings and Quantitative Data
Application of this protocol across yeast, plant, and mammalian cell models consistently demonstrated that impairment of the glutathione system caused a much more severe disruption of NADP redox homeostasis during oxidative challenge than did inhibition of the thioredoxin system [3]. This revealed the unexpected and conserved role for the glutathione system as the primary mediator of antioxidative electron flux [3].
Table 2: Summary of Key Experimental Findings on Antioxidant Pathway Contributions
| Experimental Model | Intervention | Observed Effect on NADP Redox State | Interpretation |
|---|---|---|---|
| Yeast, Mammalian, Plant Cells [3] | Glutathione (GSH) depletion (DNCB) | Severe oxidation & poor recovery after H₂O₂ challenge | Glutathione system is primary electron mediator |
| Yeast, Mammalian, Plant Cells [3] | Thioredoxin inhibition (Auranofin) | Moderate impact on NADP redox state after H₂O₂ | Thioredoxin system plays a secondary role |
| Isolated Heart Mitochondria [62] | Combined GSH depletion & Trx inhibition | Synergistic increase in H₂O₂ emission (6-17 fold) | Systems act concertedly; GSH handles basal load |
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Reagents for NADP(H) Biosensor Research
| Reagent / Tool | Function / Description | Example Use in Research |
|---|---|---|
| NAPstar Biosensors | Genetically encoded, ratiometric sensors for NADPH/NADP+ ratio | Real-time monitoring of subcellular NADP redox state in live cells [3] |
| Auranofin (AF) | Selective inhibitor of Thioredoxin Reductase (TrxR) | Probing the contribution of the thioredoxin system to antioxidant defense [3] [62] |
| 2,4-Dinitrochlorobenzene (DNCB) | Electrophilic agent that depletes cellular glutathione (GSH) | Investigating the role of the glutathione system in redox homeostasis [3] [62] |
| Peredox | Ratiometric biosensor for NADH/NAD+ redox state | Monitoring cellular energy metabolism and NAD(H) dynamics [30] |
| SoNar & iNap | High dynamic range sensors for NAD(H) and NADPH respectively | Tracking fluctuations in NADH/NAD+ ratio or NADPH concentration [30] |
| H₂O₂ | Reactive oxygen species (ROS) inducer | Applying controlled oxidative stress to challenge cellular antioxidant systems [3] |
The discovery of the glutathione system's primacy exemplifies the power of genetically encoded biosensors like NAPstars to uncover fundamental biological principles. These tools provide a window into the dynamic and compartmentalized world of cellular metabolism, moving beyond static snapshots to real-time observation. The experimental protocols outlined here provide a framework for researchers to investigate NADP(H) dynamics in their own systems. Integrating these biosensors with targeted pharmacological inhibitors and precise environmental controls allows for the dissection of complex metabolic pathways. This approach is not only critical for basic science but also has profound implications for drug development, particularly in areas where redox imbalance is a key factor, such as in metabolic diseases, cancer, and aging-related disorders. The ability to dynamically monitor the NADPH/NADP+ balance in living cells opens new avenues for screening compounds that modulate redox homeostasis for therapeutic benefit.
Conclusion
The development and application of genetically encoded biosensors represent a transformative advancement for the dynamic regulation of the NADPH/NADP+ balance. Moving beyond static engineering strategies, these tools provide unprecedented, real-time insight into subcellular redox dynamics across diverse organisms, from bacteria to human cells. As demonstrated by platforms like NAPstars, biosensors are not merely measurement devices but are powerful engines for discovery and optimization, enabling the identification of key metabolic bottlenecks and the engineering of robust production strains. The future of this field lies in the continued expansion of biosensor specificity and reliability, their deeper integration with omics technologies and synthetic genetic circuits, and their application in elucidating the role of redox metabolism in disease mechanisms. For researchers and drug developers, embracing these biosensor-driven approaches will be crucial for unlocking new biomanufacturing pathways and pioneering novel redox-based therapeutic interventions.
References
- 1. NADPH homeostasis in cancer: functions, mechanisms ... [https://www.nature.com/articles/s41392-020-00326-0]
- 2. NAD(H) and NADP(H) Redox Couples and Cellular Energy ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5737637/]
- 3. A family of NADPH/NADP+ biosensors reveals in vivo ... [https://www.nature.com/articles/s41467-024-55302-x]
- 4. The roles of NADPH and isocitrate dehydrogenase in ... [https://www.sciencedirect.com/science/article/abs/pii/S0378595522002271]
- 5. Antioxidant Protection of NADPH-Depleted Oligodendrocyte ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4962338/]
- 6. Quantitative flux analysis reveals folate-dependent NADPH ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4104482/]
- 7. Elevating cytosolic NADPH metabolism in endothelial cells ... [https://www.nature.com/articles/s41467-025-64652-z]
- 8. Dynamic and Static Regulation of Nicotinamide Adenine ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11314019/]
- 9. Genetically encoded fluorescent sensors reveal dynamic ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5555402/]
- 10. Sources and implications of NADH/NAD+ redox imbalance in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4869616/]
- 11. Combating oxidative stress in vascular disease: NADPH ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3361719/]
- 12. NADH reductive stress drives metabolic reprogramming [https://www.sciencedirect.com/science/article/pii/S0962892425001576]
- 13. Oxidative Stress Fundamentals [https://www.mdpi.com/2076-3417/15/18/10191]
- 14. Monitoring NAD(H) and NADP(H) dynamics during ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8807777/]
- 15. Development of a lysine biosensor for the dynamic regulation ... [https://microbialcellfactories.biomedcentral.com/articles/10.1186/s12934-025-02772-3]
- 16. Genetically Encoded Fluorescent Biosensors Illuminate the ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7462118/]
- 17. Next-generation Genetically Encoded Fluorescent Biosensors ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11786609/]
- 18. Illuminating mitochondrial metabolism with genetically ... [https://www.sciencedirect.com/science/article/pii/S2590137025001323]
- 19. Genetically Encoded Biosensors Based on Fluorescent ... [https://www.mdpi.com/1424-8220/21/3/795]
- 20. A genetically encoded biosensor for in vitro and in vivo ... [https://www.sciencedirect.com/science/article/abs/pii/S0956566315305248]
- 21. A red fluorescent genetically encoded biosensor for in vivo ... [https://www.nature.com/articles/s41467-025-64484-x]
- 22. Expanding Horizons in Advancements of FRET Biosensing ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12293601/]
- 23. FRET Based Biosensor: Principle Applications Recent ... [https://www.mdpi.com/2075-4418/13/8/1375]
- 24. Exploring the landscape of FRET-based molecular sensors [https://www.sciencedirect.com/science/article/abs/pii/S0734975024001605]
- 25. Ligand recognition mechanism in genetically encoded ... [https://www.sciencedirect.com/science/article/abs/pii/S1359511325002582]
- 26. Innovations from Design to Applications in Synthetic Biology [https://www.mdpi.com/2079-6374/15/4/221]
- 27. Highly multiplexed design of an allosteric transcription ... [https://www.nature.com/articles/s41467-024-54260-8]
- 28. An intramolecular FRET biosensor for the detection of SARS ... [https://pubs.rsc.org/en/content/articlehtml/2025/nr/d4nr05040a]
- 29. Efficient Sampling of Genetically Encoded Biosensor ... [https://pubmed.ncbi.nlm.nih.gov/41182995/]
- 30. Monitoring NAD(H) and NADP(H) dynamics during organismal development with genetically encoded fluorescent biosensors | Cell Regeneration | Full Text [https://cellregeneration.springeropen.com/articles/10.1186/s13619-021-00105-4]
- 31. Semisynthetic biosensors for mapping cellular ... [https://elifesciences.org/articles/32638]
- 32. NERNST: a genetically-encoded ratiometric non-destructive sensing tool to estimate NADP(H) redox status in bacterial, plant and animal systems | Nature Communications [https://www.nature.com/articles/s41467-023-38739-4]
- 33. NADPH biosensor-based identification of an alcohol ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6969876/]
- 34. ELISA assays and NADH/NADPH conversion detection [https://www.bmglabtech.com/en/application-notes/overview-of-elisa-assays-and-nadhnadph-conversion-detection/]
- 35. Dynamic and Static Regulation of Nicotinamide Adenine ... [https://www.mdpi.com/1420-3049/29/15/3687]
- 36. Biosensor-driven strain engineering reveals key cellular ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12551699/]
- 37. Systems and synthetic metabolic engineering for amino ... [https://www.sciencedirect.com/science/article/abs/pii/S0958166911007683]
- 38. Meta-analysis of NAD(P)(H) quantification results exhibits ... [https://www.nature.com/articles/s41598-023-29607-8]
- 39. Oxidative Stress Induces the Phosphorylation of NAD+ to ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12553597/]
- 40. Researchers develop sensors for the “charge” of biological ... [https://www.uni-muenster.de/news/view.php?cmdid=14478&lang=en]
- 41. Real-time monitoring of stromal NADPH levels in ... [https://www.sciencedirect.com/science/article/abs/pii/S0981942824009288]
- 42. Physiology and Robustness of Yeasts Exposed to Dynamic ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12152528/]
- 43. Researchers develop sensors for the “charge” of biological ... [https://www.eurekalert.org/news-releases/1068904]
- 44. The role of blue light in plant stress responses [https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2025.1554281/full]
- 45. Enhancing plant resilience to environmental stressors [https://www.sciencedirect.com/science/article/abs/pii/S0885576525000098]
- 46. In Vivo NADH/NAD+ Biosensing Reveals the Dynamics of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7534465/]
- 47. Addressing the Selectivity of Enzyme Biosensors: Solutions ... [https://www.mdpi.com/1424-8220/21/9/3038]
- 48. -based identification of an alcohol dehydrogenase... NADPH biosensor [https://amb-express.springeropen.com/articles/10.1186/s13568-020-0946-7]
- 49. Addgene: A family of NADPH /NADP(+) biosensors reveals in vivo... [https://www.addgene.org/browse/article/28252672/]
- 50. Highly sensitive printed pH sensor using CB/PANI ... [https://www.sciencedirect.com/science/article/abs/pii/S095656632500867X]
- 51. Redox‐dependent binding and conformational equilibria ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12519055/]
- 52. Continuous monitoring of biomolecular targets in vivo ... [https://www.sciencedirect.com/science/article/pii/S2451910325001243]
- 53. Developing a Redox Imbalance Forces Drive (RIFD) ... [https://www.sciencedirect.com/science/article/abs/pii/S1096717625001181]
- 54. Design of a programmable biosensor-CRISPRi genetic ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6954435/]
- 55. Biosensor-assisted titratable CRISPRi high-throughput ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9845192/]
- 56. Biosensor-assisted CRISPRi high-throughput screening to ... [https://www.sciencedirect.com/science/article/pii/S2405805X24000176]
- 57. An engineered biosensor enables dynamic aspartate ... [https://elifesciences.org/reviewed-preprints/90024v2/pdf]
- 58. Decoding Molecular Network Dynamics in Cells: Advances ... [https://www.mdpi.com/2079-6374/15/9/614]
- 59. A general method for the development of multicolor ... [https://www.nature.com/articles/s41589-023-01350-1]
- 60. Genetically-Encoded Biosensors for Visualizing Live-cell ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5388356/]
- 61. Real Time Monitoring of NADPH Concentrations in ... [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2018.02564/full]
- 62. Glutathione/thioredoxin systems modulate mitochondrial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3362521/]